Tabela de conteúdos
1. Biosfera
1.1. Níveis de organização biológica
1.2. Biodiversidade e conservação de espécies
2. Biomoléculas
2.1. Biomoléculas inôrgânicas
2.2. Biomoléculas orgânicas
. 2.2.1. Ácidos nucleicos
. 2.2.2. Prótidos
. 2.2.3. Glícidos
. 2.2.4. Lípidos
3. A célula
3.1. Organitos presentes nas células
3.2. Modelo de Mosaico Fluido
3.3. Tranportes transmembranares
3.4. Importância do sistema endomembranar na digestão intracelular
4. Obtenção de matéria por seres pluricelulares
4.1. Heterotrofismo e evolução dos tubos digestivos
4.2. Autotrofismo
. 4.2.1. Fotoautotrofismo
. 4.2.2. Quimioautotrofismo
5. Sistemas de transporte em seres pluricelulares
5.1. Transporte nas plantas
5.1.1. Translocação xilémica
5.1.2. Translocação floémica
5.2. Tranporte nos animais
6. Processos de obtenção de energia
6.1. Fermentação
6.2. Respiração aeróbia
7. Trocas gasosas entre seres pluricelulares e o ambiente
7.1. Trocas gasosas em plantas
7.2. Trocas gasosas em animais
8. Regulação nervosa e hormonal
8.1. Regulação nervosa e hormonal em animais
. 8.1.1. Transmissão de mensagens nervosas
. 8.1.2. Termorregulação
. 8.1.3. Transmissão de mensagens hormonais
. 8.1.4. Osmorregulação
8.2. Regulação hormonal em plantas
9. O dogma central da Biologia
9.1. Replicação do DNA
9.2. Transcrição do DNA
9.3. Tradução do RNA
10. A vida e divisão celular
10.1. Regeneração e diferenciação celular em organismos pluricelulares
10.2. Divisão celular por meiose
11. Estratégias de reprodução
11.1. Reprodução assexuada
. 11.1.1. Clonagem artificial
11.2. Reprodução sexuada
11.3. Ciclos de vida
. 11.3.1. Ciclos de vida de seres haplontes
. 11.3.2. Ciclos de vida de seres haplodiplontes
. 11.3.3. Ciclos de vida de seres diplontes
12. Evolução biológica
12.1. Origem da célula eucarionte
12.2. Origem da multicelularidade
12.3. Origens do pensamento evolucionista
. 12.3.1. Do lamarckismo ao darwinismo e neodarwinismo
. 12.3.2. Argumentos do evolucionismo
13. Sistemática dos seres vivos
13.1. Características estruturais dos seres vivos
13.2. Classificação taxonómica actual
13.3. Classificação por domínios
1. A Biosfera
1.1. Níveis de organização biológica
Na imagem abaixo, as relações bióticas referidas são aquelas que se estabelecem entre organismos pertencentes à mesma comunidade. Geralmente, são relações tróficas (relacionadas com a obtenção de alimento), pelo que envolvem: produtores (seres autotróficos, que produzem o seu alimento através da quimiossíntese ou da fotossíntese), que constituem o primeiro nível trófico; consumidores (alimentam-se dos produtores, directa ou indirectamente), que compõem os níveis tróficos seguintes; e decompositores, que transformam a matéria orgânica em matéria inorgânica, devolvendo-a ao solo.

1.2. Biodiversidade e conservação de espécies
A biodiversidade de um ambiente é definida pelo grande número e vasta variedade de organismos vivos que nele habitam. Isto é essencial para o Homem, tanto ao nível da saúde (diferentes organismos têm diferentes compostos que podem ser usados para o diagnóstico e tratamento de doenças) como ao nível da agricultura (quanto maior a variedade de espécies, maior a probabilidade de haver sobreviventes a doenças, alterações climáticas, etc.).
Contudo, por vezes, ocorrem extinções naturais (devido a modificações naturais do clima, actividade vulcânica, etc.) – de fundo, quando uma ou mais espécies desaparecem, ou em massa, quando um grande número de espécies morre devido a catástrofes naturais globais – ou antropogénicas (quando um vasto número de espécies se extingue devido à intervenção do Homem nos ecossistemas). Para evitar estas situações, é necessário promover a conservação das espécies, identificando as comunidades em perigo e procedendo à eliminação ou neutralização dos agentes causadores do seu declínio (com a criação de parques e reservas cuja legislação protege as espécies, por exemplo).
2. Biomoléculas
2.1. Biomoléculas inorgânicas
A água é o constituinte maioritário das células, funcionando como reguladora térmica e solvente onde ocorrem inúmeras reacções químicas essenciais para o seu desenvolvimento.
Ainda que a molécula de H2O seja, como um todo, electricamente neutra, é polar, pois a sua nuvem electrónica está mais deslocada para o núcleo de um dos átomos (temos uma extremidade de carga eléctrica negativa, com mais electrões (o átomo de O), e uma extremidade de carga eléctrica positiva com menos electrões (os átomos de H)). É esta característica que permite a formação de pontes de hidrogénio entre as moléculas de água e entre estas moléculas e outras substâncias polares e contribui para o poder solvente da água.
Os sais minerais são iões inorgânicos pouco abundantes na célula, mas de grande importância. Têm funções estruturais (formação do esqueleto e dos dentes) e reguladoras (são activadores de moléculas e intervêm na actividade dos músculos e células nervosas).
2.2. Biomoléculas orgânicas
Existem quatro grandes tipos de biomoléculas nas células: prótidos, glícidos, lípidos e ácidos nucleicos. Todos eles são polímeros pelo que resultam da repetição de monómeros (= a sua unidade básica).
2.2.1. Ácidos nucleicos
Os ácidos nucleicos são substâncias intervenientes na transmissão de características genéticas. Considera-se que existem dois tipos – DNA, ou ácido desoxirribonucleico, e RNA, ou ácido ribonucleico –, ambos formados pela polimerização de nucleótidos. Estes monómeros (nucleótidos) são compostos por uma pentose (para o DNA, a desoxirribose e para o RNA a ribose) ligada (ligação éster), no carbono 5’, a um grupo fosfato, e, no carbono 3’ ao grupo fosfato (também uma ligação éster) do nucleótido seguinte (a colocação dos nucleótidos para formar os ácidos dá-se no sentido 5’→3’). O carbono 1’ da pentose está associado a uma base azotada que pode ser púrica, (de anel duplo: adenina, A, e guanina, G) ou pirimídica (anel simples: timina, T, uracilo, U, ou citosina, C), and this is where the plot thickens, visto que, enquanto no DNA só se encontram nucléotidos contendo bases A, T, C e G, no RNA, só existem U, A, G e C. Analisemos, por isso, cada ácido individualmente.
 Uma molécula de DNA é formada por duas cadeias polinucleotídicas antiparalelas (o princípio de uma, no carbono 5’ do último nucleótido, corresponde ao fim da outra, ou seja, à sua extremidade 3’) enroladas em dupla hélice e ligadas entre si por pontes de hidrogénio que se estabelecem entre suas bases azotadas (A, T, C e G), de acordo com o princípio da complementaridade: os nucleótidos com A unem-se sempre aos T por duas pontes (escreve-se A=T) e os C aos G com três destas ligações (escreve-se C≡G). O princípio da complementaridade de bases, que impede que a molécula de DNA tenha largura variável e seja, por isso, quimicamente instável, permite-nos, como apontou Chargaff, escrever: (A+G)/(T+C)=1.
Uma molécula de DNA é formada por duas cadeias polinucleotídicas antiparalelas (o princípio de uma, no carbono 5’ do último nucleótido, corresponde ao fim da outra, ou seja, à sua extremidade 3’) enroladas em dupla hélice e ligadas entre si por pontes de hidrogénio que se estabelecem entre suas bases azotadas (A, T, C e G), de acordo com o princípio da complementaridade: os nucleótidos com A unem-se sempre aos T por duas pontes (escreve-se A=T) e os C aos G com três destas ligações (escreve-se C≡G). O princípio da complementaridade de bases, que impede que a molécula de DNA tenha largura variável e seja, por isso, quimicamente instável, permite-nos, como apontou Chargaff, escrever: (A+G)/(T+C)=1. Sendo o DNA um composto universal e transversal à maior parte das espécies, as variações que podemos encontrar entre estas moléculas no mundo biológico são no seu comprimento e número existente nas células, mas, principalmente, ao nível das sequências nucleotídicas que contém. É o genoma, conjunto dos genes (sequências de nucleótidos que correspondem a dadas características genéticas do indivíduo), que se expressa no genótipo (conjunto das informações hereditárias, em si, contidas no genoma) e afecta o fenótipo (características por si exibidas e que resultam tanto do genótipo como das condições ambientais) do ser, permitindo a distinção dos diversos indivíduos.
Nas células eucarióticas, as moléculas de DNA enrolam-se sobre si próprias e sobre várias proteínas (histonas, entre outras), originando uma substância denominada cromatina que se encontra circunscrita ao núcleo (devido às suas dimensões, não consegue atravessar os poros do invólucro nuclear) e, durante a divisão celular, condensa para originar cromossomas.


Uma molécula de RNA é formada por uma única cadeia polinucleotídica, em que as bases azotadas são A, U (o uracilo substitui a timina, sendo complementar de A), C e G, que se forma no núcleo e migra, daí, para o citoplasma das células. Pode apresentar-se sob três formas distintas:
- RNA mensageiro (mRNA) é uma molécula longa que se forma a partir da maturação da molécula originada pela transcrição do DNA e transporta a informação genética até ao local de síntese proteica. Depois de cumprida a sua função, degrada-se facilmente.
- RNA de transferência (tRNA) é uma cadeia curta interveniente na síntese proteica e na qual cerca de metade dos nucleótidos se ligam por complementaridade de bases (recorde-se que, para o RNA, A=U e C≡G) e os restantes originam três ansas: uma delas estabelece a ligação com o ribossoma, outra liga-se a uma enzima que catalisará a reacção de síntese e a restante contém uma sequência de três nucleótidos (anticodão) complementares de outros três (codão) do mRNA. A extremidade 3’ liga-se a um aminoácido, transportando-o consigo até ao local da síntese proteica.
- RNA ribossómico (rRNA) é uma molécula larga e dobrada que se enrola sobre algumas proteínas para formar as unidades do ribossoma que coordenará a interacção entre os tRNA e o mRNA que intervêm na síntese de uma proteína.
2.2.1. Prótidos
Os prótidos são compostos quaternários, ou seja, têm, obrigatoriamente, quatro tipos de átomos: de carbono (C), de hidrogénio (H), de oxigénio (O) e de azoto (N). De acordo com a sua complexidade, podem ser classificados como aminoácidos, péptidos e proteínas.
 Os aminoácidos são a unidade básica dos prótidos. Possuem um átomo central de carbono ligado a grupo amina, a um grupo carboxilo, a um átomo de hidrogénio e a um radical (porção da molécula que varia conforme o tipo de aminoácido).
Os aminoácidos são a unidade básica dos prótidos. Possuem um átomo central de carbono ligado a grupo amina, a um grupo carboxilo, a um átomo de hidrogénio e a um radical (porção da molécula que varia conforme o tipo de aminoácido). Os péptidos são cadeias de aminoácidos unidos por uma ligação peptídica, estabelecida entre o grupo carboxilo de um aminoácido e o grupo amina do seguinte. Chamamos oligopéptidos às cadeias compostas por 2 a 20 aminoácidos (quando são formadas por 2, também podemos designá-las por dipéptidos; com 3, tripéptidos; and so on, and so forth) e polipéptidos às que ultrapassam esse número.


As proteínas são compostas por uma ou mais cadeias polipéptidicas e têm uma estrutura tridimensional definida que nos permite classificá-las em níveis de organização: estrutura primária (sequência de aminoácidos), secundária (sequência de aminoácidos em hélice ou folha pregueada, devido à formação de pontes de hidrogénio entre os compostos), terciária (proteína de estrutura secundária que se dobra sobre si própria, ficando com um aspecto globular) ou quaternária (várias cadeias de estrutura terciária que estabelecem ligações entre si). Têm várias funções: função de transporte, estrutural, reguladora (fazem parte de algumas hormonas), imunológica (de defesa), de armazenamento, contráctil (são o componente maioritário dos músculos) e enzimática (são catalisadores de reacções químicas).

2.2.3. Glícidos
Os glícidos ou hidratos de carbono são compostos ternários, ou seja, têm, obrigatoriamente, três tipos de átomos: de carbono (C), de hidrogénio (H) e de oxigénio (O). Estas moléculas têm funções energéticas (como o amido e glicogénio, que são polissacarídeos de reserva energética vegetal e animal, respectivamente) e estruturais (como a celulose (na parede celular vegetal), a quitina (carapaça dos insectos) e a heparina (anticoagulante)). De acordo com a sua complexidade, podem ser classificados como monossacarídeos, oligossacarídeos ou polissacarídeos.
Os monossacarídeos, ou oses, são a unidade básica dos glícidos e, embora possam ser de cadeia aberta, em solução aquosa dispõe-se sempre em anel. A sua fórmula química é dada por (CH2O)n, em que n é o número de átomos de carbono da molécula (entre 3 e 9), e podem ter cadeia aberta ou fechada (anel). Assim, podem ser classificados em trioses (n=3), tetroses (n=4), pentoses (n=5), etc.
Os oligossacarídeos são cadeias lineares (como a celulose) ou ramificadas (como a amilopectina) com entre 2 e 10 monossacarídeos, unidos por uma ligação glicosídica. A partir deste número, designam-se polissacarídeos.
2.2.4. Lípidos
Os lípidos são um grupo cujas moléculas, constituídas por carbono (C), hidrogénio (H) oxigénio (O) e outros átomos, são insolúveis e água e solúveis em solventes orgânicos apolares (como benzeno, por exemplo). Podem ter funções energéticas (glicerídios), estruturais (como os fosfolípidos e as ceras) e reguladoras (esteróides).
- Os glicerídios são compostos formados por ácidos gordos saturados (cadeias de átomos de carbono ligados entre si por ligações simples, ligados a átomos de hidrogénio e com um grupo final carboxilo) ou insaturados (cadeias de átomos de carbono ligados entre si por ligações duplas ou triplas, ligados a átomos de hidrogénio e com um grupo final carboxilo) e glicerol (álcool com três grupos hidroxilo), unidos por uma ligação éster. O nome da molécula depende do número de ácidos gordos a que se alia o glicerol: monoglicerídeo (um ácido gordo), diglicerídeo (dois ácidos gordos) ou triglicerídeo (três ácidos gordos). Os mais comuns nos seres vivos são as gorduras (células animais) e os óleos (células vegetais).
- Os fosfolípidos são compostos formados por um grupo variável ligado a um grupo fosfato que está unido a um diglicerídeo. São moléculas anfipáticas (têm uma extremidade polar e hidrofílica e uma extremidade apolar e hidrofóbica) e as principais constituintes da membrana citoplasmática.
- As ceras são compostos formados pela união de um ácido gordo de cadeia longa com um álcool de cadeia longa. Funcionam como material impermeabilizante de folhas, frutos, penugem das aves, etc.
- Os esteróides são compostos formados por ácidos gordos e álcoois de cadeia cíclica contribuem para a formação das hormonas esteróides e componentes da bílis.
3. A célula
De acordo com a Teoria Celular, a célula é a unidade básica de reprodução, desenvolvimento e hereditariedade do ser vivo, só podendo formar-se a partir de outra pré-existente.
3.1. Organitos presentes nas células

3.2. Modelo de Mosaico Fluido
A membrana celular ou plasmalema é um invólucro que separa o meio intracelular do extracelular. Segundo o modelo actualmente aceite (Modelo de Mosaico Fluido), celular é composta por lípidos, proteínas e alguns glícidos.
A uma dupla camada de fosfolípidos (em que as extremidades apolares de cada camada estão unidas), cujas moléculas podem realizar movimentos laterais, transversais (flip-flop), de flexão ou de rotação, que, em conjunto com o colesterol que se lhes une, conferem à membrana uma grande fluidez, podem estar associadas a proteínas intrínsecas ou integradas (ligadas fortemente à extremidades apolares dos fosfolípidos), extrínsecas ou periféricas (estão à superfície e unidas à membrana por ligações frágeis) ou transmembranares (integradas que atravessam a toda a membrana). Encontramos, ainda, alguns glícidos, que, ao associarem-se aos fosfolípidos (glicolípidos) ou às proteínas (glicoproteínas), formam o glicocálix, cuja função é o reconhecimento de certas substâncias por parte da célula.

3.3. Transportes transmembranares
A osmose é o transporte passivo (= sem consumo de energia) de água através das aquaporinas da membrana plasmática de acordo com o gradiente de concentrações em água (do meio com menor concentração em soluto (meio hipotónico) e maior em água para o meio com maior concentração em soluto (meio hipertónico) e menor em água). Este movimento dá-se por fenómenos físicos, sendo a sua velocidade proporcional à diferença de concentrações. Quando a célula se encontra inserida num meio hipotónico (solução aquosa com concentração em soluto menor à do interior da célula), entrará água na célula, o que fará com que esta inche (fica túrgida, isto é, num estado de turgescência) ou rebente (lise celular). Pelo contrário, num meio hipertónico (solução aquosa com concentração em soluto maior à do interior da célula), haverá um fluxo de água para o exterior da célula e esta encolherá (fica plasmolisada, isto é, num estado de plasmólise). Num meio isotónico (solução aquosa com concentração em soluto igual à do interior da célula), a célula permanecerá num estado normal.

A difusão simples é o transporte passivo (= sem consumo de energia) de outras substâncias (como alguns iões e moléculas apolares) através da membrana plasmática de acordo com o gradiente de concentrações (do meio com maior concentração na substância para o meio com menor concentração). Este movimento dá-se por fenómenos físicos, sendo a sua velocidade proporcional à diferença de concentrações.
A difusão facilitada é o transporte passivo (= sem consumo de energia) de algumas moléculas polares de dimensões consideráveis (glicose, aminoácidos, vitaminas…) através da membrana plasmática de acordo com o gradiente de concentrações (do meio com maior concentração na substância para o meio com menor concentração) com a intervenção de permeases. As substâncias ligam-se à parte hidrofílica destas proteínas que, ao modificarem a sua forma, as expulsam para o interior da célula. Este movimento não se dá por fenómenos físicos. A velocidade da difusão facilitada aumenta quando a diferença de concentrações aumenta, mas só até ao ponto de saturação das permeases, quando estabiliza.

O transporte activo é o transporte de substâncias necessárias ao equilíbrio da célula através de proteínas transportadoras e contra o gradiente de concentrações (isto é, do meio com menor concentração na substância para o meio com maior concentração). Este movimento não se dá por fenómenos físicos e requer gasto da energia resultante da hidrólise ATP (adenosina trifosfato). A bomba de sódio e potássio é um exemplo muito estudado. Às proteínas mediadoras (ATPases) ligam-se três iões de sódio e uma molécula de ATP, que, ao dividir-se em Pi (que fica ligado à ATPase) e ADP (adenosina difosfato), fornece à proteína energia para que esta mude a sua forma. Os três iões são libertados e dois iões de potássio ligam-se à ATPase, que liberta o Pi (que se junta ao ADP, formando nova molécula de ADP). A proteína retoma a sua forma inicial, libertando os iões de potássio e o processo recomeça.

A endocitose é o transporte de material para o interior da célula por invaginação da membrana plasmática. Pode ser de três tipos: fagocitose (a célula emite pseudópodes e engloba a partícula numa vesícula fagocítica), pinocitose (entram líquidos ou soluções na célula numa vesícula endocítica) ou endocitose mediada por receptor (em que as macromoléculas se ligam às vesículas endocíticas que as conduzem até ao interior da célula.
A exocitose é o transporte de material para o exterior da célula através de vesículas de secreção que se fundem com a membrana plasmática.
3.4. Importância do sistema endomembranar na digestão intracelular
A digestão intracelular é realizada com a intervenção do sistema endomembranar, isto é, do conjunto da membrana citoplasmática com o retículo endoplasmático (RE), o complexo de Golgi (conjunto de dictiossomas) e os lisossomas.
O retículo endoplasmático está intimamente ligado à membrana nuclear e pode ser liso (REL) ou rugoso (RER). Este último possui vários ribossomas ligados à sua face, pelo que, quando estes produzem proteínas, o retículo transporta estas moléculas até a um dos dictiossomas da célula (por canalículos ou envolvendo-as e formando uma vesícula de transporte que se fundirá com a face de formação do dictiossoma). Aí, estas substâncias são transformadas em proteínas funcionais ou enzimas. Caso sejam formadas enzimas, o dictiossoma expulsa estas substâncias através de um lisossoma (que é, no fundo, apenas uma cisterna que se desprende da face de maturação deste organelo). Unindo-se com vesículas endocíticas, estas estruturas formam um vacúolo digestivo e digerem as substâncias captadas por endocitose (heterofagia). Quando se unem as vesículas que contêm organelos celulares que precisam de ser substituídos, formam um vacúolo autofágico e digerem-nas (autofagia).
Os resíduos da digestão (= substâncias de que a célula não necessita) são transportados até ao meio extracelular por uma vesícula de secreção que se funde com a membrana citoplasmática.

4. Obtenção de matéria por seres pluricelulares
4.1. Heterotrofismo e evolução dos tubos digestivos
Os seres heterotróficos obtêm, geralmente, a matéria necessária para o seu metabolismo em três fases: ingestão, que consiste na introdução da matéria no organismo; digestão, que é o conjunto de processos mecânicos (através de órgãos como os dentes, por exemplo) e químicos (por acção de enzimas que provocam reacções de hidrólise) que permitem a transformação das macromoléculas dos alimentos em moléculas mais simples; e absorção, que designa a passagem das moléculas simples resultantes da digestão para o meio interno do ser vivo. Nestes casos, a digestão é intracorporal (ocorre dentro do organismo), podendo ser intracelular ou extracelular.
Quando a digestão é extracorporal, não ocorre ingestão de matéria, sendo que a obtenção de matéria ocorre apenas em duas fases: digestão (fora do corpo do organismo) e absorção.
A digestão intracorporal apresenta mais vantagens para o ser vivo do que a digestão extracorporal, pois, uma vez que esta ocorre dentro do corpo, as moléculas simples que dela resultam (pela transformação de macromoléculas) já se encontram no organismo (ao contrário do que sucede na digestão extracelular), pelo que a sua absorção é mais eficaz. De entre os dois tipos de digestão intracorporal (extracelular e intracelular), o mais vantajoso é a digestão extracelular, pois permite aos seres vivos ingerir e armazenar grandes quantidades de matéria que é lentamente digerida, não sendo, por isso, necessário, ingerir continuamente alimento (como sucede nos organismos que realizam digestão intracelular). Examinemos, pois, a digestão intracorporal e como ela se processa em quatro animais distintos: a hidra, a planária, a minhoca e o ser humano.
- A hidra de água doce é um organismo com um tubo digestivo incompleto (= tem uma só abertura) denominado cavidade gastrovascular (que é, simultaneamente, um órgão digestivo e um órgão de distribuição). O alimento capturado entra nesta cavidade através da boca e é digerido pelas enzimas libertadas pelas células glandulares existentes na sua parede (digestão extracelular). Os produtos resultantes são fagocitados pelas restantes células da cavidade, sendo novamente digeridos em vacúolos digestivos e distribuídos pelo citoplasma da célula e pelas restantes células (digestão intracelular). Ocorre, então, digestão extracelular e digestão intracelular, sendo que os produtos não aproveitados são expulsos novamente pela boca.
- A planária também possui um tubo digestivo com uma só abertura (= incompleto), mas a cavidade digestiva já apresenta alguma diferenciação, sendo possível distinguir três órgãos: boca, faringe (que se projecta da primeira) e intestino (onde ocorre a digestão e absorção). Este último órgão possui ramificações que garantem uma maior área de absorção e que, por isso, conferem maior eficácia ao processo da digestão. Pode, então, dizer-se que o sistema digestivo da planária é mais evoluído do que o da hidra. Contudo, tal como sucede com a hidra, os resíduos da digestão são expelidos pela boca.
- A minhoca possui um tubo digestivo completo (= com duas aberturas), com vários órgãos especializados. O alimento é ingerido através da boca, passando pela faringe e pelo esófago, e é armazenado no estômago. A digestão dá-se na moela, onde é o alimento triturado (transformações mecânicas) e, em seguida, no intestino, onde sofre a acção das enzimas (transformações químicas). A passagem dos produtos resultantes para os vasos sanguíneos ocorre, também, no intestino, cuja prega dorsal — a tiflosole— aumenta a área de absorção, aumentando a sua eficácia. Os resíduos eliminam-se pelo ânus, pelo que o fluxo de matéria no seu tubo digestivo se processa numa só direcção. Assim, este sistema digestivo é mais evoluído do que o da planária.
- O ser humano possui um tubo digestivo com duas aberturas (= completo): boca e ânus. Os alimentos são ingeridos pela boca e é nela que se inicia a digestão através da acção da enzima amílase e da língua e dentes. O bolo alimentar aí formado segue, pelo esófago, para o estômago, onde é novamente sujeito a enzimas, mas também a movimentos peristálticos e a ácido clorídrico. Forma-se, assim, o quimo, que segue para o intestino delgado onde, no duodeno, é misturado com os sucos intestinal e pancreático (que contêm outras enzimas) e com a bílis, sendo convertido em quilo. A existência de pregas contendo vilosidades formadas por células com microvilosidades no intestino delgado aumenta a área de absorção das moléculas simples, tornando-a mais eficaz. Os produtos a eliminar seguem para o intestino grosso (onde é absorvida alguma água) e são expelidos pelo ânus. Este sistema, tal como o da minhoca, assegura que o fluxo de matéria é unidireccional. Contudo, visto os seus órgãos serem mais especializados (com destaque para o intestino, que, na minhoca, só tinha uma prega), é mais evoluído que o deste anelídeo (hidra < planária < minhoca < Homem).
Podemos concluir que os tubos digestivos completos representam um passo evolutivo face aos incompletos pois o facto de possuírem duas aberturas faz com que a matéria siga uma só direcção, o que permite ao ser vivo ingerir, digerir, absorver e expelir produtos em simultâneo. (Os organismos com tubos digestivos incompletos não podem ingerir e expelir produtos ao mesmo tempo, pois há apenas uma abertura.) Por outro lado, a existência de pregas ou ramificações no tubo aumenta a área de absorção, tornando o processo mais eficaz.
4.2. Autotrofismo
4.2.1. Fotoautotrofismo
Durante a fase fotoquímica, ao incidir luz sobre um organismo que realiza fotossíntese, as moléculas de água presentes no estroma dos cloroplastos das suas células são decompostas em dois electrões, dois iões H+ e um átomo de oxigénio (fotólise). Simultaneamente, os electrões dos pigmentos fotossintéticos dos fotossistemas II (P680) e I (P700) das membranas dos tilacóides dos cloroplastos das suas células captam a energia luminosa, ficando excitados. Assim, os seus electrões vão "saltando" de pigmento em pigmento, ficando o que o perde oxidado e o que o recebe reduzido, até chegarem ao centro da reacção (um par de clorofilas a). Do P680, os dois electrões das clorofilas são captados por uma molécula aceptora, que os conduz através uma cadeia transportadora constituída por proteínas que recolhe a energia despendida pelas partículas durante as reacções de oxirredução ao longo dessa estrutura e a aproveita para fosforilar o ADP, convertendo-o em ATP. (De acordo com a hipótese quimiosmótica, a energia é, na verdade, utilizada para bombear os H+ resultantes da fotólise da água para o interior do tilacóide, donde, atravessando ATPases, são transportadas passivamente para o estroma, e provocam alterações nessas proteínas que unem os ADP a grupos Pi.) Do P700, os dois electrões são transportados por uma cadeia semelhante até uma molécula de NADP+ que, ao juntarem-se a um ião H+ (resultante da fotólise da água), reduzem, convertendo-a em NADPH. Os electrões da água repõem os perdidos pelo P680, completando-se fotofosforilação acíclica.
Pelo processo cima descrito, o organismo produz quantidades iguais de ATP e de NADPH; porém, uma vez que este necessita de mais ATP do que NADPH para realizar a fotossíntese, realiza, por vezes, um outro tipo de fotofosforilação: a fotofosforilação cíclica. Esta só envolve o P700, cujos electrões, ao serem excitados pela luz, vão "saltando" de pigmento em pigmento, ficando o que o perde oxidado e o que o recebe reduzido, até chegarem ao centro da reacção (um par de clorofilas a). As clorofilas cedem os electrões a uma molécula aceptora (que fica, então, reduzida, enquanto que as clorofilas ficam oxidadas) que os conduz a uma cadeia transportadora constituída por proteínas. A energia dos e- é captada pela cadeia e utilizada para fosforilar o ADP (convertendo-o em ATP) e estes são reconduzidos ao P700. E recomeça o ciclo.
Os seres fotoautotróficos aproveitam igualmente todos os comprimentos de onda da luz? Não. Estes podem conter vários pigmentos fotossintéticos, que se dividem em três categorias: clorofilas (a, b, c e d), carotenóides (carotenos e xantófilas) ou ficobilinas (ficobilinas e ficoeritrinas). Cada um deles tem uma determinada cor, o que significa que, ao ser atingido pela luz, reflecte essa dada cor (= esse comprimento de onda), absorvendo todas as outras (ou seja, todos os outros comprimentos de onda). Física básica. Ora, a única radiação que é usada pela planta é a que é absorvida por estes pigmentos e, embora os seres fotoautotróficos tenham vários com várias cores nos seus cloroplastos, a sua concentração não é igual, o que significa que haverá comprimentos de onda que não são ou quase não são aproveitados (não há pigmentos suficientes para absorver uma quantidade significativa desta radiação).
Terminada a fase fotoquímica, a planta avança para a fase química ou não dependente directamente da luz que consiste no Ciclo de Calvin, realizado no estroma do cloroplasto. Numa fase de fixação do carbono, cada molécula de CO2 é combinada, com uma molécula de RuDP (ribulose difosfato), formando um composto com seis átomos de carbono que, sendo muito instável, divide-se rapidamente em duas moléculas com três carbonos cada (PGA). A fase de produção de compostos orgânicos consiste na fosforilação do PGA pela molécula de ATP (é gasta uma molécula de ATP por cada molécula de PGA) e posterior redução da molécula resultante por acção do NADPH (é gasta uma molécula de NADPH por cada molécula a reduzir). O composto que se forma é o PGAL, ou aldeído fosfoglicérico.
Duas em cada doze moléculas de PGAL são convertidas noutros compostos orgânicos (são necessários 2 PGAL para originar uma molécula de glicose) e as restantes dez são utilizadas na fase seguinte do Ciclo de Calvin: a regeneração da RuDP. Estas dez moléculas são convertidas em seis moléculas de RuMP (ribulose monofosfato), que, ao interagirem com o ATP (gasta-se uma molécula de ATP por cada molécula de RuMP), formam seis moléculas de ribulose difosfato (RuDP), o composto que se combinará com o CO2 no recomeço do ciclo.

4.2.2. Quimioautotrofismo
A quimiossíntese é um processo de autotrofia alternativo à fotossíntese em que a produção de matéria orgânica se dá sem intervenção da luz. A fonte de energia reside reacções de oxidação de compostos minerais reduzidos (como amoníaco, dióxido de carbono e sulfureto de hidrogénio). Estas permitem a obtenção de protões e electrões excitados, sendo que as partículas são utilizadas para formar NADPH (juntando a uma molécula de NADP+ um protão e dois electrões) e a energia destas é aproveitada para formar ATP (juntando a uma molécula de ADP um grupo PI).
Numa segunda fase, ocorre o ciclo das pentoses, durante o qual, tal como no ciclo de Calvin, se produzem compostos orgânicos a partir do dióxido de carbono absorvido e com intervenção do NADPH e do ATP criados na primeira fase.
5. Sistemas de transporte em seres pluricelulares
5.1. Transporte nas plantas
5.1.1. Translocação xilémica

1. A hipótese da pressão radicular sugere que a ascensão da seiva bruta no xilema se deve à criação de forças osmóticas ao nível da raiz. Os iões presentes no solo são transportados para as células dos pêlos radiculares das raízes (que existem para aumentar a área de absorção) da planta por transporte activo dos iões presentes no solo para o seu interior (quando estes estão em maior quantidade no meio intracelular) ou difusão simples (quando estes estão em maior quantidade no meio extracelular). Como consequência, estas células ficam hipertónicas em relação ao meio exterior, pelo que a entrada de água nestas se faz por osmose. A acumulação da água nestes tecidos cria uma pressão que "empurra" a água (que leva consigo alguns sais), através do xilema, até às folhas. Esta teoria é apoiada pela exsudação caulinar (saída contínua de seiva bruta através de um corte no caule de uma planta) e pela gutação (saída de água pelos hidátodos, com consequente formação de pequenas gotas nas margens das folhas). Contudo, visto que a pressão radicular medida em algumas plantas (em particular, as de grande porte) não explicaria a ascensão da seixa xilémica, que a maioria das plantas não apresenta gutação ou exsudação (havendo, até, algumas que absorvem água na zona dos cortes em vez de a perderem) e que há plantas em que não se verifica qualquer pressão radicular, foi necessário encontrar uma nova explicação.


2. A hipótese da tensão-coesão-adesão relaciona a transpiração com a absorção radicular. A subida da temperatura causa a abertura dos estomas, localizados nas folhas. A consequente transpiração (perda de água na forma de vapor) cria uma força de tensão ou sucção, devido ao défice de água criado. A sucção estende-se até à raiz, que absorverá água cuja subida será facilitada pela coesão entre as suas moléculas (estão ligadas por pontes de hidrogénio, formando uma coluna) e pela adesão destas aos vasos xilémicos, fenómenos que se devem à polaridade da molécula de H2O. Sendo a transpiração provocada pelas altas temperaturas a causa da absorção de água, pode dizer-se que a energia solar (que aquece a Terra) é o motor da ascensão xilémica. Assim, durante o dia, quando a temperatura é mais alta, os estomas abrem e a quantidade de água absorvida é maior e, durante a noite, quando a temperatura é mais baixa, os estomas fecham e a quantidade de água absorvida é maior.


5.1.2. Translocação floémica
O floema é, na planta, o tecido que faz o transporte da seiva elaborada ou floémica, isto é, da água e compostos orgânicos, desde as folhas até ao resto da planta. É constituído por quatro tipos de células: células dos tubos crivosos, que são alongadas e estão vivas (embora tenham perdido a maior parte dos seus organitos) e ligadas topo a topo, sendo que a união (placas crivosas) apresenta inúmeras perfurações; células de companhia, que estão vivas, possuem todos os organelos de uma célula normal e se localizam junto das células dos tubos crivosos, com as quais mantém ligações citoplasmáticas, ajudando-as no seu funcionamento; fibras, que são alongadas e, apesar de mortas, têm funções de suporte; e células do parênquima, vivas e pouco diferenciadas, que desempenham funções de reserva.

A hipótese do fluxo de massa admite que a translocação floémica se deve a um gradiente de concentração em sacarose entre o local de produção e o local de consumo ou armaze-namento. As células da folha, que realizam a fotossíntese, produzem glícidos (glicose e frutose) que serão convertidos em sacarose. Este produto passa de célula em célula até atingir as células de companhia do floema e, depois, as células dos tubos crivosos, onde se acumula. A passagem da sacarose das células de companhia para as células dos tubos crivosos é feita por transporte activo porque as últimas possuem uma maior concentração neste composto (devido à sua acumulação aí) do que as primeiras. A maior concentração em sacarose das células dos tubos crivosos vai, também, fazer com que estas sejam meios hipertónicos, havendo, por isso, entrada de água por osmose das células em redor (incluindo o xilema). Assim, a célula fica túrgida (aumenta a pressão de turgescência), o que faz com que o seu conteúdo (água e sacarose) seja "empurrado" através das placas crivosas para a célula do tubo crivoso seguinte. A sacarose sai, depois, por difusão simples, da célula do tubo crivoso para as células de companhia para chegar a um local de armazenamento ou consumo e, quando tal sucede, a água em excesso também abandona a célula para as células em redor (incluindo o xilema).
Esta teoria é confirmada pelas experiências realizadas em afídeos, insectos que parasitam a planta e que, ao penetrarem os caules com os seus aparelhos bucais, conseguem extrair a seiva floémica sem realizarem qualquer movimento de sucção devido à elevada pressão a que esta se encontra.
5.2. Transporte nos animais
Uma vez que os animais são seres heterotróficos e que, por este motivo, os seus órgãos e tecidos não sintetizam os seus próprios nutrientes, o seu sistema de transporte deve assegurar um fornecimento contínuo destas moléculas às células. Por outro lado, a constante produção de resíduos por parte dos tecidos leva à evolução do sistema de transporte, que é, por isso, nos animais, capaz de transportar mais rapidamente grandes quantidades de substâncias do que nas plantas.
Nos animais mais simples e aquáticos, não existe um sistema de transporte especializado. A hidra, pertencente ao grupo dos Cnidários, é um exemplo disto mesmo. É formada por apenas duas camadas de células, pelo que, ao entrar água na cavidade gastrovascular através da boca, todas as suas células ficam em contacto directo com o meio ambiente (note-se que a camada externa também está em contacto com a água visto a hidra estar mergulhada nesta), podendo, assim, haver difusão dos gases de e para as células, dos nutrientes para as células e dos resíduos para a água.
Nos animais mais complexos, existe um sistema de transporte composto por órgãos especializados. Estes, denominados sistemas circulatórios, apresentam, geralmente: um fluido circulante, que transporta substâncias de e para as células, sendo responsável pelo controlo da composição físico-química do meio interno, e pode apresentar ou não células especializadas (como leucócitos, hemácias, etc.) e pigmentos respiratórios (como a hemoglobina, por exemplo); um órgão propulsor, que bombeia o fluido circulante para o resto do corpo e pode estar ou não dividido em aurículas (cavidades com paredes finas para recepção do fluido) e ventrículos (cavidades com paredes grossas e musculosas que enviam o fluido para fora do órgão); e um conjunto de vasos (artérias, veias e capilares) ou espaços (seios ou lacunas) por onde circula o fluido circulante.
Os sistemas circulatórios abertos são aqueles em que o fluido circulante realiza parte do seu trajecto em vasos especializados e parte em lacunas, chamando-se, visto não haver distinção entre o sangue (líquido que circula em vasos) e a linfa (líquido que circula em lacunas, banhando os tecidos), hemolinfa. Os insectos possuem um sistema de transporte deste tipo. Um coração tubular contrai-se e impulsiona a hemolinfa através de válvulas para a aorta dorsal. Daí segue para as lacunas do seu corpo (cujo conjunto, cheio com a hemolinfa, se designa hemocélio), efectuando trocas de substâncias (nutrientes, resíduos e hormonas, mas não gases) com as células, e volta a entrar no coração pelos ostíolos (aberturas) quando este relaxa, pois, ao fazê-lo, cria uma força de sucção. O movimento da hemolinfa é, também, facilitado pelas contracções musculares do insecto.

Os sistemas circulatórios fechados são aqueles em que o fluido circulante realiza todo o seu trajecto em vasos especializados, chamando-se sangue. Nos sistemas circulatórios fechados, as trocas entre o sangue e as células é feita ao nível dos capilares sanguíneos e tendo por intermediária a linfa intersticiali (= linfa que banha os tecidos). Os anelídeos possuem um sistema deste tipo. Um vaso dorsal funciona como coração, bombeando o sangue, com as suas contracções, para os arcos aórticos da sua parte interior, que o enviarão para um vaso ventral. Daqui, o sangue parte de regresso ao coração, atravessando uma rede de capilares que irriga os tecidos e culmina no vaso dorsal.

Nos peixes, o sangue que aflui ao coração é venoso e entra, atravessando o seio venoso, para a sua única aurícula. Segue para o seu único ventrículo e daí, atravessando o cone arterial, para as brânquias, onde é oxigenado (durante a hematose branquial) e, portanto, convertido em sangue arterial. Depois, é recolhido na artéria aorta, que se ramifica em arteríolas e, por fim, em capilares, conduzindo o sangue aos tecidos. Os capilares reúnem-se em vénulas, que, por sua vez, se reúnem em veias, levando o fluido de volta ao coração. Visto que o sangue só atravessa o coração uma vez por circulação, a circulação dos peixes é simples.
Nos anfíbios, o sangue venoso entra para a aurícula direita do coração, através do seio venoso. Segue para o seu único ventrículo e daí, atravessando o cone arterial, para os pulmões e para a pele, onde é oxigenado (durante a hematose pulmonar e cutânea, respectivamente) e, portanto, convertido, em sangue arterial. Regressa ao coração pela aurícula esquerda, reúne-se novamente no ventrículo, onde se mistura parcialmente com o sangue venoso, e segue, atravessando o cone arterial, para os tecidos pela artéria aorta. Visto que o sangue atravessa o coração duas vezes por circulação (realiza duas circulações: a pulmonar e a sistémica), a circulação é dupla, mas, como há mistura parcial dos sangues venoso e arterial, é, simultaneamente, incompleta.
Nos répteis, a circulação é realizada de modo semelhante à dos anfíbios, mas a mistura de sangue é minimizada, também, pela existência de um septo incompleto a dividir o único ventrículo do coração. Tal como nos anfíbios, a circulação é dupla e incompleta.
Nas aves e mamíferos, o sangue venoso entra no coração, através das veias cavas, para a aurícula direita. Segue para o ventrículo direito através da válvula bicúspide, que o envia, através da válvula semi-lunar direita, para a artéria pulmonar. Esta guia-o até aos pulmões, onde este é oxigenado (durante a hematose pulmonar) e donde é levado, pela veia pulmonar, de volta ao coração. Entra pela aurícula esquerda, atravessa a válvula tricúspide para o ventrículo esquerdo e é bombeado, através da válvula semi-lunaresquerda para a artéria aorta, que o conduz a todos os tecidos do corpo. Visto que o sangue atravessa o coração duas vezes por circulação (realiza duas circulações: a pulmonar e a sistémica), a circulação das aves e mamíferos é dupla e, como não há mistura dos sangues venoso e arterial, é, simultaneamente, completa.

De uma perspectiva evolutiva, sistema circulatório aberto < sistema circulatório fechado, porque a deslocação do fluido é mais eficaz quando é feita dentro de vasos concebidos para o efeito, já que, quando esse é o caso, a velocidade de circulação é maior. Por outro lado, estando na presença de sistemas fechados, temos circulação simples < circulação dupla, já que, quando o sangue regressa ao coração depois de ser convertido em sangue arterial, é bombeado para o resto do corpo com uma maior pressão, aumentando-se a eficácia de oxigenação dos tecidos. No caso de circulação dupla, circulação incompleta < circulação completa porque as misturas de sangue arterial e venoso reduzem a concentração em oxigénio do sangue que aflui aos tecidos e, consequentemente, reduzem a eficácia da sua oxigenação. O facto das células dos mamíferos e aves receberem sangue mais oxigenado e com maior pressão que as dos répteis ou anfíbios, faz com que apresentem uma maior capacidade energética e permite a homeotermia (= manutenção de uma temperatura corporal constante). Logo, sistema circulatório aberto < sistema circulatório fechado com circulação simples < sistema circulatório fechado com circulação dupla e incompleta < sistema circulatório fechado com circulação dupla e completa.
6. Processos de obtenção de energia
O metabolismo celular traduz-se em reacções de catabolismo (degradação) e de anabolismo (síntese). As primeiras conduzem à libertação de energia, podendo, pois, distinguir-se três métodos de produção desta: respiração aeróbia (aceptor final é o oxigénio), respiração anaeróbia (aceptor final é uma molécula inorgânica) e fermentação (aceptor final é uma molécula orgânica).
6.1. Fermentação
A fermentação é um processo de obtenção de energia a partir da molécula de glicose (C6H12O6) realizado em condições anaeróbias (= sem oxigénio). Ocorre no hialoplasma (parte do citoplasma) das células, em duas etapas: glicólise (dividida em fase de activação e fase de rendimento) e redução do ácido pirúvico.
A molécula de glicose é quimicamente inactiva, pelo que, para que a sua degradação se inicie, tem que ser activada. Assim, é fosforilada por duas moléculas de ATP, convertendo em frutose-difosfato (fase de activação da glicólise). Este composto divide-se imediatamente em duas moléculas de aldeído fosfoglicérico e cada uma delas é oxidada por NAD+ (formando-se, assim, NADH + H+ e sendo a energia resultante deste processo canalizada para a formação, a partir de dois ADP e de dois grupo Pi, de duas molécula de ATP), transformando-se em ácido pirúvico (fase de rendimento da glicólise). Assim, durante a glicólise, formam-se 2 NADH + 2 H+, duas moléculas de ácido pirúvico e 4 ATP e gastam-se 2 ATP (saldo energético = 4 - 2 = 2 ATP).

A etapa de redução do ácido pirúvico (ou redução do piruvato) pode conduzir à formação de diferentes produtos. Durante a fermentação alcoólica, ocorre a descarboxilação (liberta-se dióxido de carbono) das duas moléculas de ácido pirúvico seguida da sua redução por acção do NADH (gasta-se uma molécula destas por cada molécula de ácido pirúvico), sintetizando-se, assim, duas moléculas de álcool etílico (uma por cada molécula de ácido pirúvico). Durante a fermentação láctica, as duas moléculas de ácido pirúvico são reduzidas por acção do NADH (gasta-se uma molécula destas por cada molécula de ácido pirúvico), sintetizando-se duas moléculas de ácido láctico (uma por cada molécula de ácido pirúvico).

6.2. Respiração aeróbia
A respiração aeróbia é um processo de obtenção de energia a partir da molécula de glicose (C6H12O6) realizado em condições aeróbias (= com oxigénio). Compreende quatro etapas: glicólise (ocorre no hialoplasma), formação de acetil-coenzima A (na matriz mitocondrial), ciclo de Krebs (na matriz mitocondrial) e cadeia respiratória (na membrana interna da mitocôndria).
A etapa da glicólise processa-se do mesmo modo que na fermentação. Por cada molécula de glicose, temos, no fim desta fase, 2 ATP, 2 NADH e 2 moléculas de ácido pirúvico. Na presença de oxigénio, as duas moléculas de ácido pirúvico entram na mitocôndria, onde são descarboxiladas (cada uma perde um carbono, que, juntando-se ao O2, forma uma molécula de dióxido de carbono) e oxidadas (reduzindo, assim, cada uma delas, um NAD+ a NADH + H+), transformando-se em duas moléculas de acetil. Estas moléculas unem-se, cada uma, a uma coenzima A, concluindo-se a segunda etapa da respiração aeróbia. Então, por cada molécula de glicose, temos, no fim desta fase, 2 ATP, 4 NADH, 2 acetil-CoA e 2 moléculas de dióxido de carbono libertadas.
O ciclo de Krebs ou do ácido cítrico inicia-se com a libertação, por parte de um acetil-CoA, da sua coenzima A (esta tem, por isso, apenas um papel de transporte). Ao interagir com o ácido oxaloacético (4 carbonos), o acetil (2 carbonos) origina ácido cítrico (6 carbonos), que é descarboxilado (perde 2 carbonos, formando 2 moléculas de CO2) e oxidado (forma 3 NADH e reduz um FAD a FADH2). A energia destas reacções é canalizada para a produção de uma molécula de ATP a partir de ADP e de um grupo Pi, sendo que o composto resultante é o ácido oxaloacético (4 carbonos) com que se inicia este ciclo. Então, por cada molécula de glicose, temos, no fim desta fase, 4 ATP, 10 NADH, 2 FADH2 e 6 moléculas de dióxido de carbono libertadas.

Os electrões acumulados pelos 10 NADH e 2 FADH2 são cedidos, depois, a proteínas aceptoras que os transmitem a outras proteínas seguintes (pertencentes à cadeia respiratória) numa sequência de reacções redox, ou de oxidação-redução (a proteína que cede a partícula fica oxidada e a que a recebe é reduzida), até serem captados pelo aceptor final: o oxigénio. A energia resultante destas é utilizada para formar ATP a partir de ADP e de um grupo Pi, sendo que se formam 3 destas moléculas por cada NADH e 2 por cada FADH2 (o valor é menor para o segundo composto porque este cede as suas partículas a uma proteína a meio da cadeia, não havendo um aproveitamento tão grande da sua energia). Como 3*10+2*2=34, por cada molécula de glicose, temos, no fim desta fase, água, 38 ATP e 6 moléculas de dióxido de carbono libertadas.

7. Trocas gasosas entre seres pluricelulares e o ambiente
7.1. Trocas gasosas em plantas
As trocas gasosas (e transpiração) das plantas realizam-se através de estomas, estruturas que se localizam, principalmente, na superfície inferior das folhas (embora também existam em menor quantidade noutras regiões das plantas e são formadas por duas células-guarda especializadas que delimitam um orifício – o ostíolo. Estas células, que, à semelhança das da epiderme, realizam fotossíntese, possuem certas características distintivas: semelhantes a um feijão e com paredes celulares de espessura assimétrica (a parede em contacto com a epiderme foliar é mais fina), as células são cobertas por microfibrilas de celulose.
São vários os factores que levam à abertura ou fecho do estoma: a quantidade de dióxido de carbono do ar e a luz (visto que estes são essencial para a fotossíntese, o estoma abrir-se-á se a concentração de CO2 e a luminosidade forem grandes), o pH do meio (num meio ácido, os estomas fecham), a humidade do solo e do ar (uma planta em stress hídrico fechará os seus estomas, por onde se dá a transpiração) ou mesmo o próprio ritmo biológico do ser poderão variar o tamanho do ostíolo. Quando as condições são favoráveis, pelo transporte activo de iões de potássio para as células-guarda, instala-se uma pressão osmótica (as células tornam-se um meio hipertónico em relação à epiderme) que leva à entrada de água nestas. Ficando túrgidas, as células-guarda deformam-se na região com parede mais fina, abrindo o ostíolo e, por conseguinte, o estoma. O fecho dos estomas é provocado pela diminuição da pressão de turgescência após difusão dos catiões de potássio para fora das células-guarda.

7.2. Trocas gasosas em animais
A respiração celular, nos animais, traduz-se nas trocas gasosas que ocorrem entre estes e o ambiente por fenómenos de difusão gasosa ao longo de superfícies respiratórias com quatro características comuns: são pouco espessas, permitindo o contacto eficaz com o meio; apresentam-se húmidas, o que facilita a dissolução e difusão dos gases, são altamente vascularizadas (caso o animal possua um sistema de transporte diferenciado), para facilitar o contacto com o fluido circulante; garantem uma grande área de contacto com os meios interno e externo devido ao desenvolvimento de protuberâncias, por exemplo, de função semelhante às das vilosidades intestinais.
Em animais muito simples, como os cnidários, os platelmintes e os nematelmintes, que possuem somente uma ou duas camadas de células, as trocas gasosas realizam-se através da superfície corporal e directamente entre o meio intracelular e o ambiente exterior ao organismo: por exemplo, nas hidras, as camadas exterior e interior de células difundem e absorvem gases directamente do meio aquático e da cavidade gastrovascular do animal, respectivamente, enquanto a forma achatada das planárias e seres semelhantes confere às suas células uma grande facilidade na troca gasosa directa. Em seres mais complexos, as trocas gasosas através da superfície corporal ou tegumento, ocorrem em seres cuja pele é húmida e pode, por isso, servir de meio de passagem de gases para o sistema circulatório. É o caso dos anelídeos, mas também de alguns anfíbios e peixes, que realizam este tipo de hematose conjugado com outro tipo de trocas gasosas.

Nos peixes, a hematose é branquial, dando-se através das brânquias ou guelras, formadas por evaginações da superfície corporal e em contacto directo com a água. Em peixes ósseos, estras estruturas são compostas por séries de filamentos branquiais altamente vascularizados (possuem uma artéria aferente e outra eferente unidas por uma rede capilar numa dilatação do filamento chamada lamela) inseridos em arcos branqueais e estão protegidas, na cavidade branquial, pelo opérculo. A água entra pela boca e atravessa as lamelas, oxigenando o sangue que flui na rede de capilares que aí se encontra e libertando o dióxido de carbono deste, sendo a eficácia da troca gasosa assegurada pelo mecanismo de contracorrente (o fluxo de água tem sentido contrário do do sangue na rede capilar branquial, permitindo uma maior taxa de difusão de gases) dos peixes.

Os insectos e outros artrópodes terrestres respiram através da entrada e saída de ar por orifícios (espiráculos) em traqueias que, ao ramificarem-se em redes de traquíolas, tubos em contacto directo com as células e que executam a troca gasosa (não há intervenção do sistema circulatório, pelo que a difusão é directa). As traqueias mantêm-se abertas graças a uma estrutura quitinizada em forma de hélice e, em algumas espécies, válvulas controlam a entrada de ar.

Para além da hematose cutânea (= hematose através da superfície corporal), os anfíbios realizam trocas gasosas através da superfície de pulmões bastante simples e com uma área de superfície pequena antecedidos por uma laringo-tranqueia. Os répteis, mais adaptados à vida terrestre, possuem uma traqueia ramificada em brônquios ligados a pulmões mais complexos e nos quais se diferenciam alguns sacos alveolares que aumentam a área da superfície de troca gasosa. Os mamíferos apresentam pulmões grandes constituídos por inúmeros sacos alveolares que estão unidos a bronquíolos (ramificações dos dois brônquios em que se divide a traqueia) e são altamente vascularizados, circulando o sangue lentamente nessa região, de modo a permitir a difusão de gases. Apesar das diferenças nos sistemas respiratórios, tanto os anfíbios como os répteis e os mamíferos efectuam as trocas gasosas em dois momentos – o de inspiração (o ar entra no sistema e viaja até aos pulmões, onde se dá a troca gasosa) e expiração (o ar, rico em CO2, abandona os pulmões pelo percurso inverso) – que não podem, por isso, ocorrer em simultâneo.

De facto, a vantagem evolucionária do sistema respiratório das aves em relação ao dos mamíferos (grupo, dos supracitados, com pulmões mais desenvolvidos) reside no facto do primeiro permitir aos voador executar esses dois movimentos (de inspiração e expiração) simultaneamente. O ar entra, pela traqueia, nos sacos aéreos posteriores, onde é armazenado e, após nova inspiração (e expiração), empurrado até a um par de pulmões pequenos onde, em parabrônquios, ocorre uma troca de gases cuja eficácia é maximizada por um mecanismo de contracorrente semelhante ao dos peixes (nos parabrônquios, o ar circula no sentido oposto ao do fluxo sanguíneo). Outra entrada/saída de ar conduz o ar rico em CO2 até aos sacos aéreos anteriores, onde permanece até que ocorra uma nova inspiração.
Independentemente do método de troca gasosa, quando esta ocorre tendo o fluido circulante como mediador, isto é, quando é uma hematose, a respiração deve-se à diferença entre as pressões parciais dos diversos gases no sangue e o no ar/água inspirado/ingerido, pois, visto a troca constituir uma difusão, procura anular o gradiente e restaurar a igualdade entre os meios interno e externo.
8. Regulação nervosa e hormonal
Todos os seres vivos são sistemas abertos, pelo que, para evitar a perda da homeostasia, ou seja, do equilíbrio biológico (= estabilidade de condições dos órgãos do ser), face a alterações ambientais, possuem sistemas reguladores do meio interno. Na maioria dos casos, a regulação faz-se por retroalimentação negativa (o sistema procura reduzir ou contrariar a alteração), embora também possam verificar-se alguns exemplos de retroalimentação positiva que, uma vez que consistem numa amplificação, por parte do sistema, da alteração sentida, não asseguram a manutenção da homeostasia.
8.1. Regulação nervosa e hormonal em animais
8.1.1. Transmissão de mensagens nervosas
Nos animais, as regulações são feitas, em grande parte, pelo sistema nervoso, composto por dois tipos de células: os neurónios e as células-glia.
- As células glia ocupam a região entre os neurónios, desempenhando, no sistema nervoso, funções de suporte, revestimento e isolamento dessas unidades.
- Os neurónios são a unidade básica do sistema nervoso e apresentam um corpo celular com dois tipos de prolongamentos citoplasmáticos: as dendrites, prolongamentos finos e ramificados que recebem e conduzem os estímulos até ao corpo celular, e o axónio, uma fibra fina e longa que transmite o estímulo ao neurónio seguinte. Em Vertebrados e Invertebrados complexos, as fibras nervosas que, em conjunto com o tecido conjuntivo, formam os nervos, são constituídas pelo axónio e por células especializadas – as células de Schwann – que o cobrem, formando uma bainha de mielina (aumenta a velocidade de propagação dos estímulosii) segmentada (às porções de axónio desprotegido dá-se o nome de nódulos de Ranvier).
Quando recebem estímulos do meio, os nervos sensitivos (aferentes) convertem-nos em impulsos nervosos que se transmitem, ao longo da fibra nervosa, até ao sistema nervoso central. Após processamento, o SNC reenvia instruções acerca do comportamento a adoptar aos nervos motores (eferentes) que o controlam sob a forma de um novo impulso. Analisemos, pois, o seu método de propagação.
Devido, principalmente, à distribuição dos catiões de K+ e Na+ mantida, os neurónios apresentam uma diferente quantidade de cargas eléctricas negativas no meio intracelular do que as que existem no exterior, pelo existe uma diferença de potencial, denominada potencial de membrana, entre as duas faces da membrana. Quando não é atravessado por impulsos, o potencial de membrana de um neurónio é um potencial de repouso (-70 milivolts, pois a superfície interna é negativa face à externa), sendo mantido por bombas de sódio e potássio que contrariam os mecanismos de difusão e graças ao fecho dos canais de K+ e Na+ que existem ao longo da membrana.
 Porém, quando é atingindo por um estímuloiii, os canais de sódio de sódio do neurónio abrem-se e, como este ião está em menor quantidade no meio intracelular que no extracelular, dá-se uma entrada rápida de Na+ na célula que leva a que esta se torne positiva em relação ao exterior, ou seja, leva a uma alteração do potencial de membrana, ou despolarização, para, aproximadamente, +35 milivolts (potencial de acção). O potencial de acção é imediatamente propagado à área seguinte do neurónio (provoca o seu desequilíbrio), mas, em cada região, quando se atinge este pico, fecham-se os canais de Na+ e abrem-se os de K+ (estes iões positivos abandonarão o neurónio) para que se altere o potencial de membrana de modo a efectuar uma repolarização, isto é, de modo a restabelecer o potencial de repouso, que, quando obtido, é acompanhado pelo fecho dos canais de potássio (a membrana regressa ao estado inicial). É a propagação do potencial de acção das dendrites para o axónio de um neurónio com as suas consequentes sucessões de despolarizações e repolarizações da membrana que constitui o impulso nervoso, que adquire, pois, um carácter eléctrico.
Porém, quando é atingindo por um estímuloiii, os canais de sódio de sódio do neurónio abrem-se e, como este ião está em menor quantidade no meio intracelular que no extracelular, dá-se uma entrada rápida de Na+ na célula que leva a que esta se torne positiva em relação ao exterior, ou seja, leva a uma alteração do potencial de membrana, ou despolarização, para, aproximadamente, +35 milivolts (potencial de acção). O potencial de acção é imediatamente propagado à área seguinte do neurónio (provoca o seu desequilíbrio), mas, em cada região, quando se atinge este pico, fecham-se os canais de Na+ e abrem-se os de K+ (estes iões positivos abandonarão o neurónio) para que se altere o potencial de membrana de modo a efectuar uma repolarização, isto é, de modo a restabelecer o potencial de repouso, que, quando obtido, é acompanhado pelo fecho dos canais de potássio (a membrana regressa ao estado inicial). É a propagação do potencial de acção das dendrites para o axónio de um neurónio com as suas consequentes sucessões de despolarizações e repolarizações da membrana que constitui o impulso nervoso, que adquire, pois, um carácter eléctrico.- Sinapses químicas. Nas terminações dos axónios, existem vesículas com neurotransmissores de diversos tipos, que são obrigadas a fundir-se, quando atravessadas por um impulso nervoso, com a membrana da terminação (membrana pré-sináptica). Libertados na fenda sináptica que separa a terminação axónica de um neurónio da dendrite do seguinte, os neurotransmissores ligar-se-ão à proteínas reguladores dos canais de sódio da membrana pós-sináptica (= da dendrite), causando a sua alteração conformacional e consequente entrada de Na+ no neurónio. Pela entrada de cargas positivas no meio intracelular, nessa célula verifica-se uma despolarização, à qual se seguirá uma repolarização: o impulso nervoso propagar-se-á ao longo de um outro neurónio.
- Sinapses químicas neuromusculares. O impulso eléctrico induz a fusão de uma vesícula contendo acetilcolina na membrana pré-sináptica e, esta substância, em contacto com o músculo, causa a sua contracção.

- Sinapses eléctricas. Nestas regiões, existem pontos de contacto entre as membranas pré e pós-sinápticas que permitem a propagação directa do potencial de acção do axónio para o neurónio seguinte. A transmissão do impulso é, deste modo, mais rápida em relação à propagação nas sinapses químicas.
A temperatura é um factor ambiental sujeito a variações constantes, pelo que, do ponto de vista da temperatura corporal, podemos distinguir animais poiquilotérmicos, cuja temperatura corporal varia com a do meio e que, por isso, dependem de fontes de calor externa (isto é, são ectotérmicos), e homeotérmicos, que possuem uma temperatura corporal constante por ajustarem a sua taxa metabólica de modo a contrariarem as alterações do meio ambiente (isto é, por serem endotérmicos).
Estudemos, pois, o último caso. Todos os mecanismos de termorregulação são controlados pela porção nervosa do hipotálamo que, quando recebe informações das células termo-sensoriais da pele que apontam para uma alteração da temperatura do meio ambiente, procura, por retroalimentação negativa, manter a temperatura corporal do corpo.
Embora geralmente eficazes, estes mecanismos só podem ser aplicados dentro de certos limites, variáveis de espécie para espécie. De facto, quando os seus limites são ultrapassados, sejam os animais poiquilotérmicos ou homeotérmicos, estes perecem, o que explica que a temperatura, condicionando a vida dos seres, seja um factor limitante.
Estudemos, pois, o último caso. Todos os mecanismos de termorregulação são controlados pela porção nervosa do hipotálamo que, quando recebe informações das células termo-sensoriais da pele que apontam para uma alteração da temperatura do meio ambiente, procura, por retroalimentação negativa, manter a temperatura corporal do corpo.
- Quando a temperatura ambiente aumenta, o hipotálamo impele o centro vasomotor do bolbo raquitidiano a provocar a dilatação dos vasos sanguíneos, de modo a aumentar o ritmo de transferência de calor para o exterior. A sudorese, visto a evaporação do suor na superfície da pele libertar energia do meio interno, e redução da taxa de metabólica, que procura minimizar a produção de calor, são outros mecanismos de reposição da temperatura normal.
- Quando a temperatura ambiente diminui, o hipotálamo impele o centro vasomotor do bolbo raquitidiano a provocar a constrição dos vasos sanguíneos, de modo a reduzir o ritmo de transferência de calor para o exterior. A erecção dos pêlos, visto a projecção destes criar uma camada isolante que retêm o calor irradiado à superfície da pela, e aumento da taxa de metabólica (verificam-se tremuras, por exemplo), que procura maximizar a produção de calor, são outros mecanismos de reposição da temperatura normal.
Embora geralmente eficazes, estes mecanismos só podem ser aplicados dentro de certos limites, variáveis de espécie para espécie. De facto, quando os seus limites são ultrapassados, sejam os animais poiquilotérmicos ou homeotérmicos, estes perecem, o que explica que a temperatura, condicionando a vida dos seres, seja um factor limitante.
8.1.3. Transmissão de mensagens hormonais
A regulação do meio interno pode dar-se, também, não graças a impulsos eléctricos mas a hormonas, mensageiros de natureza química secretados por órgãos especializados (glândulas endócrinas, como o pâncreas, que produz insulina, e os rins, que produzem adrenalina) e lançados na corrente sanguínea (actuam, pois, mais lentamente que os estímulos eléctricos) que, ao atingirem as células-alvo para os quais são designados, provocam alterações nestas de modo a mudar as condições do sistema biológico, geralmente por períodos de tempo mais duradouros que os efeitos das mensagens nervosas. O conjunto das glândulas endócrinas, que compõe o sistema endócrino, está, porém, intimamente ligado ao sistema nervoso, visto que a produção hormonal é, muitas vezes, regulada por estímulos nervosos: na verdade, a maior parte da actividade endócrina é controlada pelo hipotálamo, região do cérebro que estabelece a comunicação entre os dois sistemas ao produzir, após estimulação nervosa ou hormonal, neuro-hormonas que ou atingem o lóbulo anterior, de natureza endócrina, da hipófise e são daí lançadas como hormonas na corrente sanguínea, ou atingem o lóbulo posterior, de natureza nervosa, dessa glândula e regulam a produção e libertação de hormonas hipofisárias.

8.1.4. Osmorregulação
Uma vez que o metabolismo celular requer água e sais minerais, mas também a expulsão destes sob a forma de produtos de excreção, os seres não-osmoconformantes (isto é, aqueles que não estão em equilíbrio osmótico com o meio que os rodeiaiv e não permitem que a concentração em sais do seu corpo varie com a composição química desse local) mantém o equilíbrio entre água e sais através de um processo de osmorregulação (sendo, por isso, osmorreguladores) que se traduz em adaptações no seu sistema excretor, um sistema de órgãos que procede à remoção de substâncias tóxicas para o organismo e água em excesso em três momentos: filtração do fluido circulante (sangue ou hemolinfa), reabsorção, para a corrente sanguínea, de substâncias úteis e secreção (passagem directa de moléculas, por transporte activo, para a urina).
Os peixes ósseos marinhos constituem um exemplo de seres osmorreguladores. Estes, habitando um ambiente de elevada salinidade, constituem um meio hipotónico face à água que os rodeia e, como tal, têm tendência a perder água por osmose. Assim, para manter o equilíbrio hídrico, estes animais ingeram grandes quantidades de água, excretam sais por transporte activo em células branquiais e possuem glomérulos (redes capilares onde se dá a filtração do sangue) pouco desenvolvidos que produzem quantidades pequenas, mas com grandes concentrações de soluto, de urina. Já os peixes de água doce, que ganham facilmente água por osmose, visto o seu meio interno ser hipotónico em relação ao exterior, apresentam outro tipo de adaptações: não ingerem água, possuem células branquiais que transportam activamente sais para a corrente sanguínea e possuem glomérulos muito desenvolvidos que produzem grandes quantidades de urina muito diluída.
Os animais terrestres também necessitam de sistemas excretores adaptados à humidade do meio que habitam. Os anelídeos, por exemplo, que vivem no subsolo húmido, apresentam, em cada segmento do seu corpo, um par de metanefrídeos, estruturas tubulares altamente vascularizadas que efectuam a recolha de água e produtos de excreção através de um funil rodeado de cílios e ao longo das quais ocorre, posteriormente, reabsorção de substâncias úteis para o meio interno, antes da excreção de uma urina abundante e diluída. Já os insectos e aracnídeos, adaptados a climas mais secos, apresentam um sistema de excreção constituído por túbulos de Malpighi que absorvem substâncias directamente da hemolinfa e as conduzem para o recto, de onde, após reabsorção de grandes quantidades de água e alguns sais, são excretados como resíduos semi-secos em conjugação com as fezes.
Embora existam seres, como as aves-marinhas e as tartarugas-marinhas, que, visto ingerirem água salgada juntamente com o seu alimento, possuem glândulas que excretam activamente sais, nos Vertebrados, o órgão de excreção é o rim. No Homem e nos restantes mamíferos, o sistema urinário é formado por dois rins, dois ureteres, uma uretra e uma bexiga. A medula (zona interna) e córtex (zona externa) dos rins integram milhares de nefrónios, sendo cada um deles formado por uma cápsula de Bowman para onde os produtos de excreção são filtrados após passagem da corrente sanguínea no glomérulo, e por um tubo urinífero (dividido em tubo contornado distal, ansa de Henle e tubo contornado distal), ao longo do qual se dão fenómenos de reabsorção e secreção por transporte activo selectivo de substâncias executado pelas células das paredes do tubo e por osmose provocada pelo gradiente de concentrações criado por esses movimentos consumidores de ATP.

A concentração final da urina dependerá da permeabilidade das paredes do tubo contornado distal e do tubo colector, reguladas pelo sistema endócrino, ou, mais concretamente, pela ADH, ou hormona anti-diurética (= hormona contra a produção de urina), produzida pelo hipotálamo e libertada na corrente sanguínea pela hipófise. Se, por um lado, quando a quantidade de água no sangue desce (ou seja, quando a pressão osmótica sobe), o hipotálamo detecta a alteração e leva a hipófise a libertar ADH, conduzindo ao aumento da permeabilidade do tubo colector e, assim, a um aumento da reabsorção de água para o sangue, de modo a contrariar (retroalimentação negativa) a alteração, e diminuição do volume de urina produzido, por outro lado, quando o sangue se encontra demasiado diluído, o hipotálamo detecta a diminuição da pressão osmótica e, novamente, procura contrariá-lo inibindo a libertação, por parte da hipófise, da hormona anti-diurética e, deste modo, reduzindo a permeabilidade do tubo colector e, por conseguinte, a taxa de reabsorção da água, líquido em excesso.

8.2. Regulação hormonal em plantas
À semelhança dos animais, as plantas são sensíveis às alterações do meio externo. Assim, quando estimuladas, podem crescer na direcção desse estímulo ou na oposta (tropismos como o gravitropismo dos morangueiros) ou mover-se em função da alteração sem existir crescimento (são movimentos násticos como a fotonastia dos girassóis).
Como forma de adaptação ao meio, as angiospérmicas desenvolveram diferentes períodos de floração, relacionados com o fotoperíodo e, em particular, com o número de horas de escuridão, visto que algumas plantas só produzem flores quando são expostas ao seu período crítico de obscuridade (= número mínimo de horas de escuridão necessárias para que haja floração). Os seres vivos que apresentam estes requisitos quanto são fotoperíodo são, pois, catalogados como plantas de dia longo e noite curta, se só florescem quando o período de luminosidade é superior ao de escuridão, ou plantas de dia curto e noite longa, se sucede o inverso. Por outro lado, plantas que apresentam uma grande tolerância ao fotoperíodo são indiferentes.
Embora o desenvolvimento das plantas esteja, pois, directamente relacionado com o meio (luminosidade, humidade, concentração de CO2, etc.), a regulação do seu comportamento pode, muitas vezes, ser explicados por fitormonas (ou hormonas vegetais), substâncias orgânicas sintetizadas por células não-especializadas e transportadas pelo sistema vascular vegetal que condicionam de forma variável o crescimento das plantas e que se dividem em cinco grandes grupos: auxinas, giberalinas, citoquininas, ácidos abscísico e etileno.

Como forma de adaptação ao meio, as angiospérmicas desenvolveram diferentes períodos de floração, relacionados com o fotoperíodo e, em particular, com o número de horas de escuridão, visto que algumas plantas só produzem flores quando são expostas ao seu período crítico de obscuridade (= número mínimo de horas de escuridão necessárias para que haja floração). Os seres vivos que apresentam estes requisitos quanto são fotoperíodo são, pois, catalogados como plantas de dia longo e noite curta, se só florescem quando o período de luminosidade é superior ao de escuridão, ou plantas de dia curto e noite longa, se sucede o inverso. Por outro lado, plantas que apresentam uma grande tolerância ao fotoperíodo são indiferentes.
Embora o desenvolvimento das plantas esteja, pois, directamente relacionado com o meio (luminosidade, humidade, concentração de CO2, etc.), a regulação do seu comportamento pode, muitas vezes, ser explicados por fitormonas (ou hormonas vegetais), substâncias orgânicas sintetizadas por células não-especializadas e transportadas pelo sistema vascular vegetal que condicionam de forma variável o crescimento das plantas e que se dividem em cinco grandes grupos: auxinas, giberalinas, citoquininas, ácidos abscísico e etileno.
- As auxinas são fitormonas produzidas ao nível do meristema apical e outras regiões de intensa actividade mitótica (folhas jovens, sementes, etc.) cujo efeito sobre a planta depende largamente do órgão e da concentração em que existem: por exemplo, a concentração necessária de auxinas para promover o crescimento do caule é muito superior à precisa para provocar o desenvolvimento da raiz. Outras funções das auxinas incluem alguns tropismos, a inibição do desenvolvimento de ramos laterais e da abscisão (= queda) de folhas e frutos (evitam a desintegração dos pecíolos que unem estas estruturas à planta). Quando produzidas por sementes, as auxinas provocarão um desenvolvimento maior do pericarpo e, por conseguinte, do próprio fruto.
- Tal como as auxinas, embora de forma mais lenta, as giberalinas, produzidas em zonas de elevado crescimento, provocam o desenvolvimento de caules e sementes, para além de controlarem o uso de substâncias de reserva durante a germinação das últimas.
- Sintetizadas nas raízes e sementes, as citocinas promovem a divisão celular e o crescimento dos meristemas da planta, favorecendo o desenvolvimento de ramos laterais mas inibindo a ramificação das raízes. Outras funções incluem retardar o envelhecimento dos tecidos e, portanto, a abscisão de folhas e frutos e a germinação das sementes, e provocar a abertura dos estomas.
- O etileno produzido em tecidos maduros inibe o crescimento de raízes, caules e ramos laterais. Por outro lado, acelera o envelhecimento das células vegetais, levando à abscisão de folhas e germinação das sementes, e a maturação de frutos, em particular quando essas áreas estão doentes.
- O ácido abscísico promove a abscisão dos tecidos vegetais e acumulação de substâncias nutritivas nas sementes, mas inibe a germinação das últimas e o alongamento dos caules. Em situações de stress hídrico, provoca o fecho dos estomas.

9. O dogma central da Biologia
9.1. Replicação do DNA
O que Crick apelidou o dogma central da biologia molecular estabelece que, em seres eucariontes, a informação genética é arquivada no DNA, que se encontra circunscrito ao núcleo, e, só depois de transferida para o RNA mensageiro, mRNA, pode ser usada para sintetizar as proteínas (nos ribossomas, onde se dá a tradução da informação) que serão a expressão dos genes do indivíduo.

Para que o material genético de uma célula seja transmitido à geração seguinte, é preciso que se criem cópias do seu DNA, ou seja, é preciso que este se replique. Originalmente, propuseram-se três teorias para a replicação do DNA: conservativa (defendia que as cadeias originais se mantinham, gerando-se uma cadeia completamente nova), semiconservativa (admite que a molécula original se separa nas suas duas cadeias, às quais se juntam novos nucleótidos para formar duas moléculas de DNA de cadeias de idades distintas), e dispersiva (sugere que o DNA replicado é formado por segmentos do original alternados com outros recém-sintetizados). A experiência de Meselson e Stahl provou, finalmente, que a hipótese correcta é a replicação semiconservativa.
Antes de começar a replicar-se, uma molécula de DNA é desenrolada das proteínas e separada em duas cadeias (quebra das pontes de hidrogénio que unem as bases complementares) pela acção de várias enzimas, das quais se destacam as DNA polimerases. Depois, a RNA polimerase sintetiza uma porção de RNA, à qual se dá o nome de RNA iniciador, a partir da qual, pela ligação entre os nucleótidos complementares dos das cadeias separadas, começam a formar-se as duas novas cadeias de DNA. Estas serão anti-paralelas às cadeias originais, mas ambas crescerão no sentido 5’→3’, pelo que, enquanto uma das cadeias novas se constrói pela adição sucessiva de nucleótidos, a outra compõe-se pela acção da DNA ligase, que une os segmentos de nucleótidos (fragmentos de Okazaki) que se formam à medida que a DNA polimerase “abre” a molécula original de DNA. Concluída a replicação, o RNA iniciador é removido e substituído por nucleótidos de DNA.
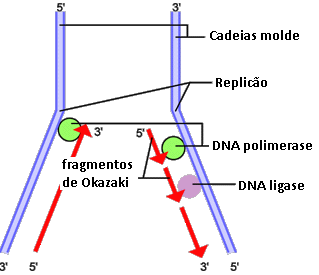
Em seres procariontes, que possuem apenas uma molécula circular de DNA, a replicação inicia-se num ponto e prossegue nas duas direcções contrárias, mas, em seres eucarióticos, o processo começa em vários locais (pontos de iniciação) do DNA, criando unidades de replicação chamadas replicões.

Por vezes, devido a agentes físicos (radiações ionizantes e ultravioletas) ou químicos (colchicina, gás mostarda, etc.), ocorrem, durante a replicação do DNA, erros que levam a que se modifique o número ou estrutura dos cromossomas de uma célula (mutação cromossomática) ou a que se altere a sequência de nucleótidos de um gene (mutação génica). Estas mutações poderão lesar o indivíduo, causando um mal funcionamento das células e, possivelmente, à sua morte, ter um impacto positivo sobre a espécie, quando se apresentam garantem ao ser características mais eficazes para o seu quotidiano e constituem um passo evolutivo, ou, quando são silenciosas, não afectar as funções normais do indivíduo de todo.
Dentro das mutações genéticas, que podem ser germinais (nos gâmetas) ou somáticas (nas restantes células), podem distinguir-se três subconjuntos: as que ocorrem por substituição de uma base do DNA (a troca pode levar à criação de tripleto que gera um codão de mRNA que codifica o mesmo aminoácido que originalmente, sendo a mutação silenciosa, ou não ter este efeito), por delecção (eliminação) de uma ou mais bases ou por inserção de bases.
9.2. Transcrição do DNA
A transcrição é um processo de cópia da informação contida no DNA para formar pré-mRNA, e dá-se em três etapas: iniciação, elongação e finalização.
- Iniciação. Quando encontra uma sequência específica de nucleótidos (o promotor, que está a montante (considera-se, para esta dicotomia, a direcção 5’→3’ da cadeia) do gene a copiar) numa região rica em nucleótidos A e T denominada TATA box, a RNA polimerase liga-se ao DNA, rompendo as pontes de hidrogénio que se estabelecem entre as bases complementares das suas duas cadeias e expondo os nucleotídeos de uma delas (aqueles que correspondem ao gene a copiar).
- Elongação. A sequência de nucleótidos exposta será o molde para a criação de RNA, que se forma, durante a elongação, pelo emparelhamento sucessivo dos nucleótidos de RNA presentes no núcleo com os de DNA de acordo com o princípio da complementaridade de bases. Enquanto esta polimerização dos nucleótidos de RNA ocorre, a RNA polimerase move-se ao longo da molécula de DNA, desenrolando-o.
- Finalização. Ao reconhecer uma zona de terminação (região rica em nucleótidos A) na molécula de DNA, uma proteína de finalização força a RNA polimerase a cessar a transcrição e a molécula de pré-mRNA formado é libertada.
Ainda no núcleo, o pré-mRNA sofre um processo de maturação durante o qual «os intrões (sequências de nucleótidos sem significado na síntese proteica) transcritos são removidos e os exões (sequências de nucleótidos que especificam aminoácidos) são ligados entre si» para formar o mRNA maduro e funcional. Porém, também pode acontecer que alguns dos exões sejam suprimidos durante o processamento do pré-mRNA (é um fenómeno chamado splicing alternativo), pelo que, consoante a combinação de exões eliminados, o mesmo gene (sequência de DNA) pode dar origem a diferentes moléculas de mRNA funcional e, como tal, a diferentes combinações de aminoácidos (ou seja, a diferentes proteínas) sintetizadas: o splicing alternativo permite ao gene expressar-se de diversas formas diferentes.

9.3. Tradução do RNA mensageiro
A tradução é o processo pelo qual a informação contida no mRNA formado durante a transcrição é descodificada pelo tRNA para dar origem a proteínas, isto é, para executar um processo de síntese proteica. Aqui, a unidade fundamental de codificação é o codão, um conjunto de três nucleótidos do mRNA que corresponde à transcição de um codogene (tripleto de nucleótidos de DNA) e ao qual se ligará um anticodão (por complementaridade de bases) presente no tRNA.
Todos os aminoácidos (a.a.) são codificados por codões, pelo que se elaborou uma tabela, o código genético, que estabelece a correspondência entre estes conjuntos de três nucleótidos de RNA e os a.a. por eles definido. Porém, embora a cada codão possa corresponder somente um aminoácido (o código é caracterizado pela sua não-ambiguidade), um aminoácido pode ser codificado por mais que uma destas unidades (a esta propriedade dá-se o nome de redundância), o que permite que o terceiro nucleótido seja o menos específico do conjunto. Alguns codões, como é o caso UAA, UAG e UGA, sendo codões de finalização (terminam a síntese proteica), não correspondem a nenhum aminoácido, mas existe um com dupla função: o codão AUG, que codifica a metionina, é o responsável pelo início da síntese proteica, sendo denominado codão de iniciação. Visto que aproximadamente todas as espécies obedecem às relações definidas no código genético, este é considerado universal.
O processo dá-se da seguinte forma:
- Iniciação. Após a saída do mRNA, através dos poros nucleares, do núcleo, este liga-se à subunidade pequena do ribossoma (localizado no retículo endoplasmático rugoso) no codão de iniciação (AUG). Aos três nucleótidos juntam-se aos do anticodão do tRNA que, pela complementaridade de bases, lhes corresponde e, por fim, a subunidade grande do ribossoma une-se ao conjunto.
- Alongamento. Enquanto o tRNA contendo o anticodão correspondente ao codão de iniciação ainda se encontra no ribossoma, um novo tRNA junta-se ao codão seguinte do RNA e o aminoácido por si transportado forma uma ligação peptídica com a metionina (aminoácido transportado pelo primeiro tRNA). Em seguida, o primeiro tRNA é libertado e o ribossoma avança três nucleótidos na cadeia de mRNA. O processo repete-se ao longo do mRNA até que se inicie a…
- Finalização. Ao chegar a um codão de finalização, o ribossoma reconhece-o pela complementaridade de bases e termina a formação de polipéptidos. A cadeia de aminoácidos, que formará uma proteína, será libertada e os componentes do complexo de tradução (mRNA, tRNAs e unidades do ribossoma) separam-se.

A síntese proteica, embora seja muito complexa, dá-se em apenas alguns minutos, sendo, por isso, dotada de grande rapidez. Devido à cópia repetida da mesma sequência de nucleótidos de DNA e à descodificação repetida das mesmas moléculas de mRNA (de facto, muitas vezes os ribossomas alinham-se, formando um polirribossoma (ver abaixo), ligados por um mRNA), pode, também dizer-se, que o processo é caracterizado pela amplificação da transcrição do DNA e tradução do RNA.

10. A vida celular
O ciclo celular, que descreve o ciclo de vida de uma célula, pode dividir-se em dois momentos: o de crescimento (interfase) e de divisão (fase mitótica). Durante a maior parte da sua vida, a célula apresenta-se em interfase, isto é, desempenhando as suas funções e em preparação para a sua divisão. Imediatamente após a sua formação, a célula inicia a síntese de proteínas (e RNA para as produzir), bem como a criação de organelos que levarão a um crescimento da célula. Após este período, denominado, G1, inicia-se o de síntese (S), no qual as moléculas de DNA do núcleo celular são replicadas, associando-se a proteínas para formar cromatina e, em células animais, forma-se um segundo par de centríolos. Assim, no momento final da interfase, G2, a célula, já tendo o seu conteúdo genético duplicado, a célula sintetiza biomoléculas (como proteínas, cuja criação requer a produção de RNA) que lhe serão úteis durante a divisão.
A fase mitótica do ciclo celular inicia-se na mitose, período de divisão do núcleo em que se distinguem quatro etapas:
- Prófase. No núcleo, a cromatina condensa para formar cromossomas constituídos por dois cromatídeos irmãos unidos por um centrómero (zona de constrição do cromossoma). Nas células animais, unindo-se por um sistema de fibrilas proteicas a que se dá o nome de fuso acromático ou mitótico (engloba as fibras do áster, microtúbulos proteicos em redor de cada centrossoma), os dois pares de centríolos formados na fase S da interfase migram para pólos opostos da célula, e, quando assumem estas posições, a membrana nuclear desorganiza-se, libertando os cromossomas para o citoplasma da célula em divisão.
- Metáfase. O material genético atinge o seu máximo de condensação, apresentando-se os cromossomas mais espessos e curtos. Em células animais, os microtúbulos proteicos do fuso acromático ligam-se aos centrómeros de cada uma destas estruturas, guiando-as para que estas se disponham ao longo de um plano equatorial (equidistante entre os pólos) da célula, com os cromatídeos apontando para os extremos desta. A disposição dos cromossomas nessa região deve-se, nas células vegetais, que não possuem centríolos, a outro tipo de microtúbulos.
- Anáfase. Após a clivagem do centrómero, separam-se os cromatídios irmãos que constituem cada um dos cromossomas, passando estes a constituir dois cromossomas independentes ligados aos microtúbulos proteicos (em células animais, do fuso acromático) por estruturas proteicas designadas cinetocoros. Em seguida, devido ao encurtamento dessas fibras, os cromossomas-filhos migram para pólos opostos da célula, num processo de ascensão polar.
- Telófase. O fuso acromático dissolve-se e, em pólos opostos da célula, os cromossomas-filhos organizam-se dentro das membranas nucleares que se formam à sua volta e descondensam, formando cromatina. No final desta fase, a célula possui dois núcleos distintos, com os seus respectivos nucléolos, em locais opostos do citoplasma.
Concluída a formação dos núcleos-filhos, o citoplasma é dividido em dois, para que se conclua a formação das novas células. Este processo, designado por citocinese, ocorre, em células animais, por estrangulamento (isto é, por constrição da membrana celular segundo o plano equatorial da célula pela contracção de um anel de filamentos aí formado), mas, nas células vegetais, cujas paredes rígidas não permitem estes movimentos, o citoplasma é separado pela fusão de vesículas denominadas fragmoplastos e derivadas do complexo de Golgi na região central da célula para originarem, após deposição de celulose, uma nova parede celular.

Para evitar a sobrevivência das células nas ocorreram erros e a transmissão destes às gerações futuras, ao longo do ciclo celular, existem diversos pontos de controlo nos quais a célula é avaliada. Os principais momentos de verificação ocorrem na interfase, em G1 e G2, e na fase mitótica: em G2, caso o DNA se encontrar muito danificado, os mecanismos de regulação do ciclo celular levam à sua estagnação naquela célula, permanecendo esta em G0 (estado de interfase, só podendo a célula dividir-se na eventualidade de receber um estímulo externo), ou à apoptose (= morte) da unidade; em G2 e fase M, os erros na replicação do DNA ou na distribuição dos cromossomas (que deverá ser equitativa), respectivamente, conduzem, geralmente, à morte celular. Quando estes mecanismos de controlo falham, podem ocorrer neoplasias malignas (cancros), que resultam da divisão descontrolada de células com deformações prejudiciais (= malignas) e podem, devido à sua capacidade de se infiltrarem em vasos linfáticos e sanguíneos, invadir tecidos de outras partes do corpo, ou seja, metastizar-se.


10.1. Regeneração e diferenciação celular em organismos pluricelulares
Uma vez que, após a mitose e citocinese, o material genético das células-filhas é uma cópia exacta do da célula que se dividiu para as originar (resulta da replicação deste e o número de cromossomas é igual, pelo que existe estabilidade genética), todas as células dos seres pluricelulares, que se desenvolvem (crescem) e regeneram os seus tecidos por sucessivas divisões mitóticas da célula-ovo, possuem o mesmo conteúdo genético; porém, à medida que aumenta a complexidade do indivíduo, verifica-se que as suas células adquirem a capacidade de se especializar numa dada função, adaptando-se, através da activação (expressão) e desactivação (não-expressão) de genes específicos, bioquímica e morfologicamente, num processo de diferenciação celular, às condições a que se sujeitam ao desempenhar o seu papel. O controlo da expressividade dos genes pode ser feito tanto ao nível da transcrição do DNA, como do processamento do pré-mRNA (splicing alternativo) e da tradução e degradação do mRNA, mas também é influenciado por factores ambientais. Muitas vezes, como reacção a novas condições ambientais, ocorre a activação de um ou mais genes que protegem uma célula madura diferenciada pela ocorrência de uma metaplasia, isto é, transformação desta noutra de um tipo diferente.
As células com capacidade de se diferenciar, a que chamamos células-tronco ou células estaminais, podem ser adultas (tecidos adultos) ou embrionárias (tecidos de embriões), e, quanto ao nível de especialização, são classificadas como: totipotentes (células, que, com todos os genes activados, podem originar qualquer tipo de tecido do indivíduo), pluripotentes (no Homem, poderão multiplicar-se e produzir todos os tecidos excluindo a placenta e anexos embrionários), multipotentes (especializadas num tipo de tecido), unipotentes (só podem originar um tipo de células) ou diferenciadas (executam uma função específica). Devido ao seu potencial para o tratamento de algumas lesões e doenças (Parkinson, Alzheimer, leucemia, etc.), as celulas-tronco são muito procuradas, podendo ser obtidas directamente do corpo humano (a medula óssea, por exemplo, produz células-tronco adultas com alguma capacidade de diferenciação), de embriões descartados e congelados em clínicas de reprodução assistida ou por clonagem terapêutica (transferência de um núcleo saudável de uma célula já diferenciada para um óvulo não-fecundado para que este se divida formando células totipotentes sempre in vitro).
As células com capacidade de se diferenciar, a que chamamos células-tronco ou células estaminais, podem ser adultas (tecidos adultos) ou embrionárias (tecidos de embriões), e, quanto ao nível de especialização, são classificadas como: totipotentes (células, que, com todos os genes activados, podem originar qualquer tipo de tecido do indivíduo), pluripotentes (no Homem, poderão multiplicar-se e produzir todos os tecidos excluindo a placenta e anexos embrionários), multipotentes (especializadas num tipo de tecido), unipotentes (só podem originar um tipo de células) ou diferenciadas (executam uma função específica). Devido ao seu potencial para o tratamento de algumas lesões e doenças (Parkinson, Alzheimer, leucemia, etc.), as celulas-tronco são muito procuradas, podendo ser obtidas directamente do corpo humano (a medula óssea, por exemplo, produz células-tronco adultas com alguma capacidade de diferenciação), de embriões descartados e congelados em clínicas de reprodução assistida ou por clonagem terapêutica (transferência de um núcleo saudável de uma célula já diferenciada para um óvulo não-fecundado para que este se divida formando células totipotentes sempre in vitro).
[scan 1]
10.2. Divisão celular por meiose
A meiose constitui um processo de divisão celular alternativo à mitose, dividindo-se em dois momentos: a divisão I e a divisão II. A primeira divisão é reducional, pois são repartidos, por dois núcleos-filhos, os cromossomas da célula original, em quatro etapas:
- Prófase I. No núcleo diplóide (com 2n cromossomas e 2n cromatídeos), a cromatina condensa e o ocorre o emparelhamento dos cromossomas homólogos (= cromossomas com os mesmo genes, mas provenientes de dois progenitores distintos) em bivalentes ou tétradas cromatídicas, com aparecimento de pontos de quiasma (= locais de cruzamento de cromatídeos) nos quais pode ocorrer crossing-over (= transferência de material genético entre cromossomas homólogos). À semelhança do que sucede na mitose, a diferenciação dos cromossomas leva à formação do fuso acromático e à desintegração do invólucro nuclear.
- Metáfase I. As fibrilas proteicas do fuso ligam-se aos centrómeros dos cromossomas de cada bivalente, dispondo os enrolamentos de cromatina, de forma aleatória, com os pontos de quiama alinhados ao longo de um plano equatorial.
- Anáfase I. As fibrilas proteicas encurtam-se, levando à ruptura dos pontos de quiasma entre cromossomas homólogos e à sua ascensão polar.
- Telófase I. Enquanto o fuso acromático se dissolve, reorganizam-se invólucros nucleares em redor dos dois conjuntos de cromossomas (já a descondensar para formar cromatina) com os quais formarão dois núcleos haplóides (= com n cromossomas cada), que, embora tenham composição genética heterogénea, são ambos formados, cada um, por 2n cromatídeos. A citocinese nem sempre sucede esta fase, visto que a telófase I conduz a uma divisão seguinte.
A segunda divisão é equacional, uma vez que, mantendo-se o número de cromossomas dos núcleos-filhos, se reduz a sua quantidade de DNA pela separação dos cromatídeos de cada condensado de cromatina. Embora exija que aos nomes de cada uma das já conhecidas 4 etapas se acrescente o numeral cardinal romano II (prófase II, metáfase II, anáfase II e telófase II), a divisão II é um processo semelhante ao da divisão mitótica que origina, a partir de cada núcleo haplóide formado na divisão I, dois núcleos, de igual ploidia (com n cromossomas cada), com n cromatídeos cada um e geneticamente idênticos.

Assim, após ocorrência de meiose, forma-se um total de quatro núcleos haplóides (= com n cromossomas) de composição genética heterogénea com n cromatídeos cada um. Neste ponto, a citocinese é obrigatória; porém, visto que a repartição do citoplasma e organitos não é sempre igual, nem sempre as quatro células-filhas são viáveis e sobrevivem após a divisão meiótica.

Contrariamente à mitose, a meiose, através dos fenómenos de recombinação genética durante o crossing-over e da disposição aleatória dos cromossomas homólogos ao longo do plano equatorial (são repartidos pelos pólos da célula aleatoriamente), criando núcleos de composição heterogénea, assegura a variabilidade genética: o último factor mencionado, actuando por si só, garante a existência de 2n combinações de genes paternos e maternos possíveis para formar uma célula.
11. Reprodução dos seres vivos
11.1 Reprodução assexuada
A continuidade da vida é assegurada pelos processos de reprodução da espécie, que podem ser assexuados ou sexuados. A reprodução assexuada é um tipo de produção de descendência na qual não ocorre fecundação de gâmetas: de facto, a prole é originada a partir das divisões mitóticas de um só progenitor ou uma das suas células, pelo que os seres filhos são geneticamente iguais a este (são clones) e, por conseguinte, idênticos entre si. A reprodução assexuada garante, deste modo, a estabilidade genética ao longo de várias gerações, o que pode, a curto prazo, ser favorável para a espécie, mas, a longo prazo, devido a inevitáveis alterações ambientais, conduzir à extinção desta pela carência de variabilidade genética entre os indivíduos que possibilite a sobrevivência de alguns espécimes. Existem vários tipos de reprodução assexuada:
- Durante a esquizogonia ou divisão múltipla, o núcleo do progenitor duplica-se sucessivamente (mitose) e cada um dos núcleos-filhos, envolvido por uma membrana celular, constitui um merozoíto que ainda habita o citoplasma do ser original. Os descendentes são libertados pelo rompimento da membrana celular do progenitor, que, até a esse momento, os protegerá de condições adversas.
- A bipartição, cissiparidade ou divisão binária consiste na separação do organismo progenitor em dois seres de dimensões e conteúdo genético idêntico, num processo simples de mitose que resulta na perda da individualidade do ser original (este, ao dividir-se, perde algumas das suas características, em particular as suas dimensões).
- Durante a gemulação ou gemiparidade, o organismo progenitor emite uma gema ou gomo geneticamente idêntica a si e, consequentemente, de características semelhantes às suas, mas de dimensões, inicialmente, inferiores às suas, que, depois de uma fase de crescimento, se separa deste, constituindo um corpo independente. Ao longo deste processo, as características do ser original mantêm-se, podendo dizer-se que ele retém a sua individualidade.
- A fragmentação é um processo de reprodução característico de seres pluricelulares, pois exige que estes se dividam em aglomerados celulares que, por regeneração de tecidos, originem organismos-filhos completos e geneticamente idênticos ao progenitor (ser fragmentado).
- A multiplicação ou propagação vegetativa é um processo de reprodução exclusivo das plantas e ocorre devido à existência, nestas, de meristemas (aglomerados de células multipotentes) em regiões como folhas, estolhos, rizomas, tubérculos e bolbos, que, por crescimento diferenciação celular, são capazes de produzir novos organismos completos. O Homem tenta reproduzir, artificialmente, a propagação vegetativa por processos como a estacaria (introdução de fragmentos caulinares ou foliares da planta no solo que desenvolverão raízes e um novo ser), mergulhia (dobragem de um ramo da planta para o enterrar no solo, onde criará raiz), alporquia (remoção de porção da casca da árvore e envolvimento dessa porção num plástico contendo terra) e enxertia (junção das superfícies cortadas de duas partes de plantas diferentes para que exista troca de material genético) por garfo, encosto ou borbulha.
- Durante a esporulação, formam-se estruturas reprodutoras denominadas esporos, que, conforme se formem, em relação ao esporângio, interior (processo endogénico, em que cada estrutura desenvolve a sua membrana própria) ou exteriormente (processo exogenético, na qual a membrana é formada a partir da membrana do esporângio), são classificados como, respectivamente, endosporos ou exosporos, e contêm informação para originar um novo indivíduo geneticamente idêntico ao progenitor.
- Durante a partenogénese, pela replicação do DNA de um oócito não fecundado, forma-se uma célula diplóide (com a totalidade dos cromossomas característicos da espécie) que, após sucessivas divisões mitóticas, origina um indivíduo completo e geneticamente igual ao progenitor. Esta é uma alternativa de reprodução, em períodos de escassez de indivíduos machos, por exemplo, para as espécies que geram descendência sexuadamente.
11.1.1. Clonagem artificial
Como explicado anteriormente, devido à conservação do material genética durante as divisões mitóticas, a reprodução assexuada, que assenta neste tipo de replicação de células, cria clones (seres geneticamente iguais) dos organismos progenitores. A produção artificial (clonagem) destes foi desenvolvida e aperfeiçoada em laboratórios, pelo que, actualmente, são conhecidos os mecanismos de clonagem de plantas e animais.
As plantas são clonadas, geralmente, a partir de células somáticas vegetais de um espécime saudável e de características apelativas. O tecido recolhido é desinfectado, lavado em água destilada e armazenado em recipientes contendo meios de cultura nutritivos que se apresentam hermeticamente fechados e esterilizados e se encontram em condições de humidade, temperatura e luminosidade controladas. Nos bordos dos fragmentos, surgem glóbulos de células indiferenciadas a partir dos quais se desenvolvem plântulas que, após atingirem dadas dimensões, são transferidas para vasos com solos nutritivos localizados em estufas. Este tipo de cultura in vitro, denominado micropropagação vegetativa, embora, visto requerer uma técnica e equipamento especializados e esterilizados, seja muito dispendioso e posso levar à criação de grande número de plantas doentes se a original não for saudável, produz, em regra, organismos robustos em excelente estado sanitário em larga escala e muito rapidamente.
A clonagem de animais dá-se pela inserção (muitas vezes, recorrendo a descargas eléctricas para a promover) do núcleo de uma célula diferenciada (possui o número típico de cromossomas da espécies) num oócito não fecundado de que foi extraído o núcleo. Uma vez que o núcleo diferenciado possui o mesmo material genético que a célula totipotente que o originou, este pode perder a sua especificidade, reactivando os genes não expressados, e comportar-se como um núcleo de um ovo formado por fecundação, pelo que pode, através de divisões mitóticas, originar um organismo clone daquele de que foi extraído.
11.2. Reprodução sexuada
Quanto ao número de cromossomas, podemos considerar dois tipos de células: haplóides, com n cromossomas, e diplóides, com 2n cromossomas, sendo que, por convenção, n é o número de pares de cromossomas das células diplóides da espécie. A diploidia das células haplóides é restaurada apenas aquando da fecundação, momento de fusão de gâmetas (células haplontes por definição) e, portanto, de duplicação cromossómica que, sendo característica da reprodução sexuada dos seres, origina um ovo ou zigoto. Assim, uma vez que os gâmetas são, em última análise, formados por uma divisão meióticav, a reprodução sexuada, tendo em conta a aleatoriedade da disposição de cromossomas homólogos durante a meiose que origina as células germinais e da união destas, assegura a existência de 2n x 2n = 22n combinações de genes paternos e maternos possíveis para formar o zigoto, um valor que cresce consideravelmente quando se têm em consideração os fenómenos de crossing-over da meiose, o terceiro factor de variabilidade genética da reprodução sexuada.


Nos animais, os gâmetas femininos e masculinos são produzidos nas gónadas femininas ou ovários e gónadas masculinas ou testículos, respectivamente. Em algumas espécies, os indivíduos possuem ambos os órgãos, tratando-se de seres hermafroditas suficientes, quando realizam autofecundação (unem-se os gâmetas femininos e masculinos do mesmo espécime, não existindo variabilidade genética, o que sucede frequentemente em espécies cujos indivíduos são solitários), ou insuficientes, quando há fecundação cruzada (dois seres fecundam-se um ao outro). Na maior parte das espécies animais, todavia, existe unissexualismo, só possuindo os animais um tipo de gónadas, e a fecundação é externa ou interna.
- Fecundação externa efectua-se em meio líquido e, graças às moléculas nos óvulos que só permitem a entrada de espermatozóides da mesma espécie, que estão revestidos por moléculas complementares dessas, e ao sincronismo na libertação dos gâmetas por parte dos espécimes masculino e feminino, resulta na formação de um zigoto. Embora a prole produzida seja extensa em número, os gastos energéticos são, também muito elevados, devido à baixa probabilidade de ocorrência de fecundação.
- Fecundação interna efectua-se no sistema reprodutor da fêmea após deposição, por parte do macho, dos espermatozóides neste local. Este método de fecundação permite a sobrevivência dos gâmetas dos animais terrestres, que não suportariam a dessecação que se verifica nesse meio, assegurando a continuidade dessas espécies. Por outro lado, e embora a probabilidade de ocorrência de fecundação seja mais baixa e, como tal, o dispêndio energético seja menor, também a prole é reduzida: o número de crias geradas é inferior à dezena.
No reino Plantae, os gâmetas femininos (oosferas) e masculinos (anterozóides) são produzidos em gametângios com o nome de arquegónios e anterídeos.
Por exemplo, no caso particular das plantas angiospérmicas, os óvulos (oosferas) e grãos de pólen (anterozóides) que se fundirão para formar um zigoto são gerados ao nível dos carpelos e estames, respectivamente, estruturas essas que constituem os aparelhos reprodutores feminino (contém os arquegónios) e masculino (contém os anterídeos) das flores: estas podem, pois, ser hermafroditas (possuem estames e carpelos) ou unissexuadas (ou possuem estames ou possuem carpelos). A fecundação dá-se quando, após polinização directa, se houver cruzamento dos gâmetas da mesma planta, ou cruzada, se existir troca de gâmetas com outras da mesma espécie (este cenário favorece a variabilidade genética), o pólen caído sobre o estigma desenvolve uma extensão membranar denominada tubo polínico que penetra no óvulo para formar um zigoto. Nestas situações, o ovo evoluirá para constituir as sementes que, em conjunto com o pericarpo resultante do desenvolvimento e engrossamento das paredes ováricas, compõe um fruto carnudo ou seco.


Tanto para os animais como para as plantas e outros seres, a principal vantagem da reprodução sexuada sobre a assexuada prende-se com o facto da primeira garantir, graças aos três factores supramencionados, a variabilidade genética da população. Só deste modo a espécie, possuindo indivíduos de características distintas, elevará a probabilidade de sobreviver a mudanças ambientais bruscas, ainda que com uma redução significativa do número de efectivos.
Por outro lado, como depende da ocorrência de meiose, a reprodução sexuada aumenta a probabilidade das já mencionadas mutações cromossómicas nos indivíduos pelas distribuições irregulares dos cromossomas homólogos, na divisão I, ou dos cromatídeos de cada uma destas estruturas, na divisão II.
Por outro lado, como depende da ocorrência de meiose, a reprodução sexuada aumenta a probabilidade das já mencionadas mutações cromossómicas nos indivíduos pelas distribuições irregulares dos cromossomas homólogos, na divisão I, ou dos cromatídeos de cada uma destas estruturas, na divisão II.
- Mutações estruturais ocorrem quando há delecção ou duplicação de porções do cromossoma, quando um segmento cromossómico roda 180º em relação à sua posição original no cromossoma ou quando existe transferência (translocação) de material genético entre cromossomas.
- Mutações numéricas ocorrem quando há alterações do número típico de cromossomas da espécie, podendo ser classificadas como:
- Aneuploidias, se os erros ocorrem num só par de cromossomas. Nesse par, pode existir um cromossoma a mais (trissomia), dois ou mais cromossomas extra (polissomia), um cromossoma a menos (monossomia) ou nenhum cromossoma (nulissomia). Estas mutações são melhor toleradas nos cromossomas sexuais que nos restantes, aos quais se dá o nome de autossomas.
- Poliploidias, se os erros multiplicam o total de cromossomas de um ser. Se 2n for o número de cromossomas normais para a espécie, possuindo o ser 3n cromossomas, trata-se de uma triploidia, quando tem 4n, diz-se que é uma tetraploidia, etc.
11.3. Ciclos de vida
O desenvolvimento do ovo, a transição para a vida adulta e a produção de gâmetas para originar um novo zigoto são fases da existência dos seres vivos que se sucedem, completando o seu ciclo de vida. Consoante a ploidia dominante, estes ciclos podem ser classificados como haplontes, haplodiplontes ou diplontes.
11.3.1. Ciclos haplontes

- A espirogira é uma alga verde (contém cloroplastos enrolados em hélice) haplonte, formada por filamentos constituídos por camadas de células cilíndricas dispostas topo a topo. Em condições favoráveis, o ser vivo cria réplicas de si próprio assexuadamente, por fragmentação, mas, quando o meio é mais inóspito, reproduz-se de forma sexuada. Durante o último processo, em primeiro lugar, dois filamentos de espirogira aproximam-se, formando-se protuberâncias membranares nas células de ambas as tiras que se unem, originando tubos de conjugação. Através de cada um destes canais, o conteúdo citoplasmático de cada célula (n cromossomas) de um filamento viaja até se fundir com a célula do outro (n cromossomas), num processo de conjugaçãovi que origina um zigoto com 2n cromossomas. Uma meiose pós-zigótica divide esta célula em quatro haplóides e, após degeneração de três destas (devido, possivelmente, à repartição desigual do citoplasma, como explicado anteriormente), resta uma com n cromossomas que irá, por mitoses, originar uma nova espirogira adulta.

11.3.2. Ciclos haplodiplontes

- A funária é uma briófita, isto é, uma planta avascular (= não possui tecidos xilémicos e floémicos diferenciados, efectuando o transporte de água e sais minerais para os seus tecidos por osmose e difusão, respectivamente), comum em locais que retêm a humidade. A água é, por sinal, essencial na fecundação: este processo ocorre pela deslocação dos anterozóides (n cromossomas), com auxílio dos seus flagelos, até às oosferas (n cromossomas) em meio aquático, culminando com o início da geração esporófita, ou seja, na formação de um zigoto diplóide. Este dividir-se-á mitoticamente até formar um ser esporófito adulto (2n cromossomas) que, nos seus esporângios, restaurará, por meiose pré-espórica, a haplofase graças à formação de esporos (n cromossomas). Após a libertação destas células, o esporófito morre, mas os esporos germinam e originam os seres gametófitos (n cromossomas) que produzem oosferas e anterozóides por mitoses. Nas briófitas, devido à curta duração do esporófito e à sua dependência energética do gametófito, é, na verdade, o ser haplonte que constitui a parte mais desenvolvida das plantas.

- O polipódio é uma planta vascularizada mas sem flor que habita zonas húmidas e está ligada ao solo por um rizoma. Este caule subterrâneo permite-lhe realizar reprodução assexuada por multiplicação vegetativa, mas, noutras ocasiões, a reprodução faz-se sexuadamente. Sazonalmente, surgem, na página inferior dos fetos adultos (seres esporófitos, com 2n cromossomas), soros, conjuntos de esporângios, onde, por meiose pré-espórica, se formam esporos (n cromossomas) e, por conseguinte, se dá a transição da geração esporófita para a gametófita. Ao germinarem, as estruturas originam um protalo (n cromossomas) que produz oosferas e anterozóides em arquegónios e anterídeos, respectivamente, na sua face inferior. Esta, estando em contacto com a água que se acumula no solo, assegura a deslocação dos gâmetas masculinos ao encontro dos femininos (n cromossomas cada) e a fusão destes num zigoto (2n) que marca o início da geração esporófita e levará ao desenvolvimento, na face superior do protalo, do esporófito adulto a que chamamos feto. No caso dos polipódios, contrariamente ao que sucede para as funárias, é o esporófito o ser mais desenvolvido, o que representa, já, um passo evolutivo relativamente a esses seres avasculares.

11.3.3. Ciclos de seres diplontes

- Os mamíferos são animais terrestres diplontes que se reproduzem sexuadamente. No caso do Homem, os gâmetas são formados pela ocorrência, ao nível dos órgãos reprodutores, de meioses pré-gaméticas que originam espermatozóides ou, em ovários e após degeneração de três dos núcleos haplóides formados, ovócitos. A união destas células germinativas com n cromossomas durante a fecundação restaura a diploidia, reinstaurando a fase diplóide: por divisões mitóticas sucessivas o zigoto (2n cromossomas) criará um novo adulto.
12. Evolução biológica
Devido ao seu nível inferior de complexidade e à existência de registos fósseis que o comprovam, acredita-se, actualmente, que os primeiros seres eram unicelulares e procariontes. Terá sido a partir destes que se desenvolveram células eucarióticas que, agrupando-se, formaram organismos eucariontes.
12.1. Origem da célula eucariótica
Existem três posições para explicar o surgimento da vida eucariótica: o modelo autogenético, o modelo endossimbiótico e um terceiro, e mais conciliador, ponto de vista.
De acordo com o modelo autogenético, através de invaginações da membrana citoplasmática, os organismos procariontes terão formado complexos sistemas endomembranar de cuja especialização resultaram os organitos que é hoje possível observar na célula eucariótica. Contudo, embora esta teoria pareça explicar a origem do núcleo como um fenómeno de incorporação do DNA numa vesícula membranar em formação e a natureza semelhante do invólucro nuclear, retículo endoplasmático, dictiossomas e plasmalema (componentes do sistema endomembranar), prova-se insuficiente para explicar o surgimento das mitocôndrias e cloroplastos nos moldes em que os conhecemos.

Após extensas investigações, Lynn Margulis, propôs, então, um novo modelo, segundo o qual os organitos da célula eucariótica teriam sido formados quando, após fagocitose de seres procariontes de menores dimensões, uma célula (também procariótica) de maiores dimensões estabelecera com os seres incorporados, uma relação de simbiose (daí a designação do modelo endossimbiótico) que se mantivera ao longo das gerações. Esta teoria explica a dupla membrana das mitocôndrias e dos cloroplastos (a membrana interior seria a do procarionte original e a exterior a da vesícula de fagocitose), bem como o seu tamanho (semelhante ao das bactérias) e o seu comportamento semi-independente da célula (possuindo o seu próprio DNA, semelhante ao dos actuais procariontes, os organitos replicam-se de forma autónoma, como se fossem regidos por um ciclo celular próprio, e sintetizam as proteínas de que necessitam), e é apoiada pelas actuais associações simbióticas entre eucariontes e bactérias. Porém, torna-se pouco clara nas suas explicações do desenvolvimento do sistema endomembranar dos eucariontes.

Notando as lacunas e mais-valias de cada modelo, alguns investigadores acabaram por acordar numa posição mais conciliadora: com efeito, as mitocôndrias e cloroplastos resultaram de endossimbioses com procariontes aeróbios e fotossintéticos, respectivamente, mas o núcleo e todo o sistema endomembranar foram gerados pelas invaginações membranares descritas pelo modelo autogenético.
12.2. Origem da multicelularidade
Com a multiplicação das células, tornaram-se frequentes os fenómenos de predação, pelo que se tornou vantajoso possuir maiores dimensões. O crescimento das células levantou, todavia, problemas nas trocas de substâncias com o meio, visto que o aumento do volume é acompanhado por um acréscimo significativamente menor da área da superfície. A situação resolveu-se com a agregação de células menores num volume superior, ou seja, com o aparecimento da multicelularidade.

Este passo evolutivo provou-se vantajoso, não só no domínio da competição por alimento e território e na manutenção da razão área/volume adequada, mas também devido à especialização celular a que conduziu e que permitiu uma maior eficácia no consumo energético e garantiu a manutenção da homeostasia, mesmo em alguns ambientes anteriormente hostis.
Actualmente, a verdadeira multicelularidade ocorre apenas nos seres eucariontes e consiste na interdependência estrutural de um conjunto de células associadas. Porém, existem diferentes níveis de complexidade nestas associações e, quando a especialização é muito reduzida para que o conjunto seja considerado um só organismo, pode falar-se de agregados coloniais ou colónias, se as células pertencerem à mesma espécie: é o caso de Gonium, colónia simples onde não se verifica diferenciação celular, e Volvox, em que já se verifica alguma especialização das células.
12.3. Origens do pensamento evolucionista

- A hipótese fixista da abiogénese (geração espôntanea), defendida por Aristóteles e Val Helmont, admite que os seres vivos não nascem somente pela reprodução sexuada, mas também, em certas condições específicas, a partir de matéria inerte (= não viva) que contivesse um princípio activo que despoletaria a criação de vida.
- A hipótese fixista criacionista, defendida por naturalistas como Lineu, explica a origem das espécies através da criação, das mesmas, por um ser superior, Deus. Tal como a entidade que as gerava, as formas de vida seriam perfeitas, pelo que não haveria necessidade de não conservarem as suas características.
- A hipótese fixista do catastrofismo, defendida pelo Cuvier, era suportada pela visão criacionista, mas permitia explicar a existência de fósseis correspondentes a espécies extintas: de acordo com o geólogo, por vezes, por ira divina, ocorreriam catástrofes que exterminariam formas de vida que seriam, pela acção de Deus, posteriormente substituídas por outras mais perfeitas. Entre cada extinção, as características das espécies manter-se-iam inalteráveis.
A partir do século XVIII, graças aos trabalhos do criacionista Lineu no âmbito da classificação dos seres vivos, que evidenciavam as semelhanças entre grupos distintos, e a alguns dados paleontológicos, começou a contestar-se o fixismo: surgem ideias transformistas, que sugerem que existe um ancestral comum às espécies e que estas resultam pela propagação de indivíduos seleccionados pelo ambiente. Esta perspectiva evolucionista é apoiada pela geologia da época, pois, ao esboçarem a teoria do Uniformitarismo, Hutton e Lyell sugerem, para o planeta Terra, uma idade suficientemente grande para que se tornem credíveis os fenómenos de diferenciação dos seres vivos em espécies.
12.3.1. Do lamarckismo ao darwinismo e neodarwinismo

O naturalista Lamarck é o primeiro a apresentar uma teoria evolucionista consistente e fundamentada.
- As alterações no meio ambiente provocam, nos seres vivos, uma necessidade de se adaptarem, que eles concretizam através de mudanças comportamentais.
- Os novos comportamentos consistem exercitação de alguns órgãos, mais úteis nas novas condições, em detrimento de outros e levam a que os primeiros se desenvolvam e os segundos atrofiem, de acordo com a lei do uso e do desuso.
- Pela transmissão à descendência destas características (lei da herança dos caracteres adquiridos), ganhas pelo desenvolvimento e atrofia de partes do seu corpo, os seres vivos contribuíam para a adaptação e evolução da espécie.
Actualmente, a crítica ao lamarckismo incide sobre a última lei mencionada, uma vez que, visto afectarem apenas as células somáticas do indivíduo, as características adquiridas não são transmitidas à prole.

- Devido à existência de reprodução sexuada, os seres de uma população possuem diversidade de características, ou seja, variabilidade intra-específica.
- As populações crescem para lá dos limites do meio, pelo que um grande número de indivíduos é eliminado na luta pela sobrevivência.
- Os seres com características que lhes conferem maior adaptação ao ambiente têm maior facilidade em alimentar-se e executar os processos vitais básicos que os restantes, logo sobrevivem em maior número durante a luta pela sobrevivência enquanto os menos aptos/adaptados perecem mais facilmente (selecção natural).
- Os organismos mais aptos, sobrevivendo durante mais tempo, reproduzem-se mais (há reprodução diferencial, visto que nem todos os seres se multiplicam da mesma forma) e produzem maior número de descendentes que estão, também, adaptados ao meio (supondo que este se mantém constante).
- Ao longo do tempo, o aumento do número de indivíduos adaptados ao meio leva à substituição dos indivíduos menos aptos pelos mais aptos, ou seja, à evolução da população e, consequentemente, da espécie.
As principais críticas que actualmente se apresentam ao darwinismo estão relacionadas com a explicação insuficiente da origem da variabilidade intra-específica e modo de transmissão das características favoráveis à descendência.
Com base nos dados da paleontologia, sistemática e genética, principalmente, desenvolveu-se uma teoria sintética da evolução, o neodarwinismo. Este introduz o conceito de população mendeliana, que é constituída pelo «conjunto de indivíduos que se reproduz sexuadamente e partilha um determinado conjunto de genes», sendo este o fundo genético (gene pool) da população, e considera que a evolução das espécies consiste na alteração das gene pools.
- Devido à ocorrência de mutações, à aleatoriedade da separação dos cromossomas homólogos durante a divisão I da meiose e da união de gâmetas durante a fecundação e à recombinação genética por crossing-over, os seres de uma população mendeliana possuem diversidade de genes e, consequentemente, de características, ou seja, variabilidade intra-específica.
- A selecção naturalvii actua sobre a diversidade da população mendeliana, logo os indivíduos com conjuntos de genes que lhes garantem características mais vantajosas no meio em que vivem sobrevivem durante mais tempo e em maior número que os outros, reproduzindo-se mais que estes (reprodução diferencial).
- Ao longo das gerações, certos genes vão sendo substituídos por outros numa população mendeliana, alterando-se o seu fundo genético.
Porém, as alterações no fundo genético de uma população mendeliana não são sempre causadas pelas mutações e pela selecção natural. Outros factores podem intervir na diversidade genética dos conjuntos de indivíduos:
- A reprodução diferencial acentua-se quando os indivíduos da população privilegiam certos fenótipos em relação a outros, não ocorrendo panmixia (= cruzamento ao acaso).
- A selecção artificial de indivíduos para cruzamento ajuda a fixar genes específicos.
- A migração de indivíduos em idade reprodutiva entre populações mendelianas implica fluxo de genes entre estas unidades evolutivas, pelo que pode conduzir a alterações no fundo genético de cada uma delas.
- Em populações reduzidas, ocorrem, por vezes, fenómenos de deriva genética: o fundo genético varia como resultado da eliminação ou introdução de genes de forma não selectiva mas aleatória. A deriva genética por efeito gargalo ocorre pela fixação e eliminação de genes após redução considerável do número de indivíduos (e, portante, da diversidade genética) da população. Por outro lado, quando, durante uma colonização, um grupo de seres separa-se dos restantes, pode dar origem a uma população geneticamente diferente daquela de que derivou (efeito fundador).
- Em populações de maiores dimensões, a separação de uma destas unidades e isolamento das populações-filhas poderá levar a alterações do fundo genético destas capazes de originar espécies distintas.
O quadro seguinte sintetiza algumas diferenças entre as várias teorias evolucionistas supracitadas.

O argumentos que procuram invalidar o fixismo e defender a teoria sintética da evolução provêm de diferentes áreas científicas.



Nos primórdios da humanidade, o Homem agrupava a diversidade de seres vivos que o rodeavam em função da utilidade que estes tinham para si (distinguiam-se, seres comestíveis e venenosos, por exemplo) e, devido ao seu pragmatismo, o conjunto deste tipo de dicotomias é designado por classificação prática.
Aristóteles e, mais tarde, Carl von Linné abandonaram os critérios práticos, utilizando uma classificação racional, isto é, optando por dividir os seres vivos de acordo com as suas características estruturais. Devido ao baixo número de caracteres morfológicos ou fisiológicos considerados por estes naturalistas aquando da sua agregação em categorias, este sistema é artificial. Pelo contrário, as classificações utilizadas no período pós-lineano e pré-darwiniano, graças ao acentuado desenvolvimento científico da época, baseiam-se num maior número de dados e são, por isso, naturais.
Evidentemente, visto que, até ao desenvolvimento do darwinismo, predominavam as teorias fixistas, os sistemas descritos no parágrafo anterior não consideram as relações de parentesco entre os diversos grupos de seres vivos (= não tomam em consideração o factor tempo e a evolução que a ele se associam), mas apenas as suas características fenotípicas (enganadoras, devido aos órgãos análogos que encontramos em diferentes tipos de organismo), pelo que são classificações fenéticas (ou estáticas ou horizontais), representadas por dendogramas cujo comprimento dos traços não tem significado biológico ou por chaves dicotómicas.
Foram as teorias evolucionistas que, no período pós-darwiniano, levaram à criação de novos sistemas filogenéticos de classificação (ou cladísticos ou filéticos ou evolutivos) que já consideravam o factor tempo (são, por isso, verticais) e, eram, por isso representados por cladogramas (ou árvores filogenéticas) cujos traços marcam a divergência dos grupos de seres vivos, tendo comprimentos proporcionais ao intervalo de tempo no qual ocorreu essa evolução. No fundo, uma vez que também se servem de um grande número de critérios estruturais para catalogar os organismos, as classificações filogenéticas são, também, naturais.
Assim, a Sistemática de hoje é um ramo da Biologia que compreende a taxonomia, ramo que desenvolve sistemas de classificação dos seres vivos e nomenclatura dos mesmos, e a filogenia, que estabelece relações de proximidade evolutiva entre grupos distintos de organismos.

As características de que nos servimos, actualmente, nos nossos sistemas de classificação são de diversos tipos:
Desde um sistema de classificação com dois grupos de seres vivos até um sistema com cinco, a taxonomia evoluiu significativamente.

Actualmente, utiliza-se o sistema de classificação de Whittaker modificado que agrupa os seres vivos em cinco largos grupos – os reinos Monera, Protista, Plantae, Fungi e Animalia – que se distinguem em função do nível de organização celular, tipo de nutrição e interacções nos ecossistemas dos organismos que neles figuram, como pode observar-se no seguinte quadro (retirado daqui).

O reino é o taxon (= categoria taxonómica de plural taxa) mais abrangente da classificação taxonómica hierárquica que usamos, estando subdividido em filos, que por sua vez contêm várias classes. Cada classe engloba várias ordens, que incluem várias famílias, nas quais se inserem vários géneros e é dentro destes grupos que estão as espécies. Destas sete taxa principais (reino, filo, classe, ordem, família, género e espécie)viii, todas foram criadas pelo Homem, com excepção da espécie que, definindo-se como um conjunto de indivíduos que partilha o mesmo fundo genético e pode, por isso, cruzar-se entre si, originando descendência fértil, é um grupo natural (= delimitado pela própria natureza).
De acordo com este mesmo sistema, todas as categorias taxonómicas são palavras latinas ou latinizadas, surgindo, por isso, em itálico ou sublinhadas. Em relação às famílias, deve notar-se que, geralmente, dentro do reino Animalia, todas terminam em –idae, e, nas plantas, em –aceae.
A nomenclatura das espécies destaca-se devido ao facto de ser, ao contrário das outras 6 taxa principais, que se resumem a uma só palavra, binominal: o nome da espécie inclui o nome do género, com inicial maiúscula, e o restritivo ou epíteto específico, totalmente em letras minúsculas (por exemplo, para o lobo, Canis é o género e lupus é o epíteto específico, sendo o nome da espécie Canis lupus). Similarmente, as subespécies têm uma nomenclatura trinominal que obedece à fórmula género + restritivo específico + restritivo subespecífico (por exemplo, Homo sapiens sapiens). Nomes de espécies e subespécies são, geralmente, acompanhado por uma abreviatura do nome do taxonomista que catalogou em primeiro lugar os indivíduos desse tipo e, após uma vírgula, a data de publicação da descoberta.
Em 1970, através de estudos baseados em critérios moleculares e, mais especificamente, nas sequências nucleotídicas de RNA ribossómico de vários tipos de procariontes, Carl Woese concluiu que existiam dois grupos distintos destes – arqueobactérias e eubactérias, sendo que as primeiras divergiram em primeiro lugar do ancestral comum que deu origem a ambos os grupos – o que o levou a propor uma classificação baseada em três domínios: Archaea (a que corresponde o reino Archeobacteria) Bacteria (a que corresponde o reino Eubacteria) e Eukarya (a que correspondem os reinos Protista, Plantae, Fungi e Animalia).

------------------------------------------------------------------------------------------------------------------------
 i Do sangue que chega ao nível dos tecidos, cerca de 99% do volume retorna ao sistema venoso. Os restantes 1% vão formar a linfa intersticial, que é um líquido composto por plasma e leucócitos que actua como intermediário das trocas entre o sangue e os tecidos. Este fluido é, depois, recolhido por vasos capilares linfáticos, passando a designar-se linfa circulante, que se reúnem em canais colectores até atingirem o canal torácico (que conduz a linfa de volta à corrente sanguínea através da veia subclávia esquerda) ou o canal linfático direito (que conduz a linfa de volta à corrente sanguínea através da veia subclávia esquerda). A circulação da linfa deve-se à à pressão osmótica do sangue e à pressão pela contracção dos músculos. Ao longo dos vasos linfáticos surgem os gânglios linfáticos, onde há grandes quantidades de linfócitos (um tipo de leucócitos). Resumindo, o sistema linfático (conjunto gânglios + vasos linfáticos) mantém o equilíbrio dos iões no corpo, transporta certos ácidos gordos das vilosidades intestinais para o sangue, contribui para a imunidade do organismo e assegura a entrada da linfa no sistema circulatório. ↑
i Do sangue que chega ao nível dos tecidos, cerca de 99% do volume retorna ao sistema venoso. Os restantes 1% vão formar a linfa intersticial, que é um líquido composto por plasma e leucócitos que actua como intermediário das trocas entre o sangue e os tecidos. Este fluido é, depois, recolhido por vasos capilares linfáticos, passando a designar-se linfa circulante, que se reúnem em canais colectores até atingirem o canal torácico (que conduz a linfa de volta à corrente sanguínea através da veia subclávia esquerda) ou o canal linfático direito (que conduz a linfa de volta à corrente sanguínea através da veia subclávia esquerda). A circulação da linfa deve-se à à pressão osmótica do sangue e à pressão pela contracção dos músculos. Ao longo dos vasos linfáticos surgem os gânglios linfáticos, onde há grandes quantidades de linfócitos (um tipo de leucócitos). Resumindo, o sistema linfático (conjunto gânglios + vasos linfáticos) mantém o equilíbrio dos iões no corpo, transporta certos ácidos gordos das vilosidades intestinais para o sangue, contribui para a imunidade do organismo e assegura a entrada da linfa no sistema circulatório. ↑ vii A selecção natural pode ser de dois tipos: estabilizadora, se são favorecidas as características até ali tomadas como melhores, ou evolutiva, se são favorecidas uma (evolutiva direccional) ou mais (evolutiva disruptiva) das restantes. Só o segundo tipo possibilita a formação de novas espécies, visto que o primeiro assegura a homogeneização da espécie. ↑
vii A selecção natural pode ser de dois tipos: estabilizadora, se são favorecidas as características até ali tomadas como melhores, ou evolutiva, se são favorecidas uma (evolutiva direccional) ou mais (evolutiva disruptiva) das restantes. Só o segundo tipo possibilita a formação de novas espécies, visto que o primeiro assegura a homogeneização da espécie. ↑

12.3.2. Argumentos do evolucionismo
O argumentos que procuram invalidar o fixismo e defender a teoria sintética da evolução provêm de diferentes áreas científicas.
- Anatomia Comparada. Pela comparação da morfologia de seres vivos de diversos grupos, é possível estabelecer conexões entre eles. De facto, existem três tipos de estruturas que podem ser usadas como provas do evolucionismo.
- Estruturas homólogas são órgãos, encontrados em seres vivos distintos, que, embora possam desempenhar funções diferentes, têm origem embriológica e organização semelhantes. A presença destas estruturas em indivíduos diferentes aponta para a existência de um ancestral comum a ambos que originou as espécies a que pertence por evolução divergente (= tipo de evolução que origina, a partir de um ancestral comum, devido à sujeição de populações deste a meios e, portanto, pressões selectivas diferentes, dois grupos distintos de seres).

- Estruturas análogas são órgãos, encontrados em seres vivos distintos, que, embora tenham origem embriológica e anatomia diferentes, desempenham funções semelhantes. A presença destas estruturas em indivíduos diferentes aponta para a evolução convergente (= tipo de evolução que origina, a partir de ancestrais diferentes, devido à sujeição de populações destes a meios e, portanto, pressões selectivas semelhantes, dois grupos de seres com traços idênticos) dos seus antepassados.

- Estruturas vestigiais são órgãos atrofiados que são desnecessários ao organismo que os detém e podem até não ser funcionais. A presença de estruturas deste tipo num ser permite inferir que a sua espécie possuía um ancestral com os órgãos em questão funcionais e que, sujeito um ambiente em que não existia necessidade dessas extensões corporais, as apresentou, com o passar das gerações, sucessivamente menores.
- Biogeografia. A análise das espécies nas suas respectivas áreas geográficas permite concluir que, em ambientes idênticos, estas apresentam características semelhantes, mas, em meios distintos, mesmo quando pertencem à mesma classe, apresentam adaptações diferentes. Uma vez que estas observações podem ser explicadas pela evolução convergente (sujeito a pressões selectivas iguais, os seres desenvolvem-se estruturas semelhantes) e divergente (sujeito a pressões selectivas diferentes, os seres desenvolvem-se estruturas dissemelhantes), respectivamente, a biogeografia é um saber que constitui um argumento a favor do evolucionismo.

- Paleontologia. Os argumentos paleontológicas baseiam-se em fósseis de formas já extintas e de formas intermédias.
- Os registos fósseis de formas extintas contrariam a hipótese da imutabilidade das espécies, visto que provam que o planeta foi habitado por várias vagas de organismos distintos.
- As formas intermédias, de transição ou sintéticas são fósseis que possuem características de dois ou mais grupos actuais de seres vivos e que, por esse motivo, deverão tê-los originado por evolução divergente.
- Embriologia. A semelhança entre os embriões de seres de grupos taxonómicos muito distintos e a lei da recapitulação, que defende que, durante o desenvolvimento embrionário, o ser recapitula as formas embrionárias dos seus ancestrais, pareceu constituir um argumento contra o fixismo. Porém, em anos recentes, a polémica que envolve a fiabilidade destes estudos embriológicos e a validade científica das suas conclusões (nomeadamente, da lei da recapitulação) levou a que perdessem importância para os evolucionistas.

- Citologia. De acordo com a Teoria Celular, a célula é a unidade básica estrutural e funcional da vida, sendo que a sua morfologia e tipo de organitos semelhantes em grupos distintos de seres aponta para a existência de um ancestral comum.
- Bioquímica. Todos os seres vivos são constituídos pelos mesmos tipos de biomoléculas (sais minerais, prótidos, glícidos, lípidos e ácidos nucleicos), sendo as proteínas criadas por qualquer organismo com base num código genético universal. Estas semelhanças apoiam a existência de um ancestral comum e ajudam, através de testes de análise de proteínas, hibridação de DNA e reacções imunológicas (organismos semelhantes pos-suem sistemas imunitários semelhantes) a estabelecer relações de proximidade filogenética entre várias espécies actuais.
13. Sistemática dos seres vivos
Nos primórdios da humanidade, o Homem agrupava a diversidade de seres vivos que o rodeavam em função da utilidade que estes tinham para si (distinguiam-se, seres comestíveis e venenosos, por exemplo) e, devido ao seu pragmatismo, o conjunto deste tipo de dicotomias é designado por classificação prática.
Aristóteles e, mais tarde, Carl von Linné abandonaram os critérios práticos, utilizando uma classificação racional, isto é, optando por dividir os seres vivos de acordo com as suas características estruturais. Devido ao baixo número de caracteres morfológicos ou fisiológicos considerados por estes naturalistas aquando da sua agregação em categorias, este sistema é artificial. Pelo contrário, as classificações utilizadas no período pós-lineano e pré-darwiniano, graças ao acentuado desenvolvimento científico da época, baseiam-se num maior número de dados e são, por isso, naturais.
Evidentemente, visto que, até ao desenvolvimento do darwinismo, predominavam as teorias fixistas, os sistemas descritos no parágrafo anterior não consideram as relações de parentesco entre os diversos grupos de seres vivos (= não tomam em consideração o factor tempo e a evolução que a ele se associam), mas apenas as suas características fenotípicas (enganadoras, devido aos órgãos análogos que encontramos em diferentes tipos de organismo), pelo que são classificações fenéticas (ou estáticas ou horizontais), representadas por dendogramas cujo comprimento dos traços não tem significado biológico ou por chaves dicotómicas.
Foram as teorias evolucionistas que, no período pós-darwiniano, levaram à criação de novos sistemas filogenéticos de classificação (ou cladísticos ou filéticos ou evolutivos) que já consideravam o factor tempo (são, por isso, verticais) e, eram, por isso representados por cladogramas (ou árvores filogenéticas) cujos traços marcam a divergência dos grupos de seres vivos, tendo comprimentos proporcionais ao intervalo de tempo no qual ocorreu essa evolução. No fundo, uma vez que também se servem de um grande número de critérios estruturais para catalogar os organismos, as classificações filogenéticas são, também, naturais.
Assim, a Sistemática de hoje é um ramo da Biologia que compreende a taxonomia, ramo que desenvolve sistemas de classificação dos seres vivos e nomenclatura dos mesmos, e a filogenia, que estabelece relações de proximidade evolutiva entre grupos distintos de organismos.

13.1. Características estruturais dos seres vivos
As características de que nos servimos, actualmente, nos nossos sistemas de classificação são de diversos tipos:
- Sexualidade. Os organismos que se reproduzem sexuadamente podem apresentar hermafroditismo suficiente ou insuficiente ou podem ser unissexuados e efectuar fecundação interna ou externa.
- Morfologia. As comparações com base neste critério devem ter em consideração eventuais metamorfoses (= mudanças sofridas pelo corpo durante os período pós-embrionário), o polimorfismo (por vezes, existe uma diversidade de formas que os adultos podem assumir, como é possível observar nas abelhas) e a existência de órgãos análogos noutras espécies.
- Simetria corporal. Alguns organismos são assimétricos, como as esponjas, mas outros, como o Homem, apresentam simetria bilateral (apresentam um só plano de simetria), como as estrelas-do-mar, simetria radiada (apresentam vários planos de simetria que se intersectam num eixo).
- Níveis de organização estrutural.
- Quanto à organização celular, os seres podem ser procariontes ou eucariontes, dependendo do tipo de células que lhe pertencem.
- Quanto ao número de células, os organismos podem ser unicelulares solitários ou coloniais ou podem ser multicelulares. O nível de diferenciação celular nos últimos e nas colónias de unicelulares também permite caracterizar os grupos de seres vivos.
- Tipo de nutrição. Consoante o modo como obtém a matéria orgânica de que necessitam, os seres podem ser considerados:
- Autotróficos, se produzirem matéria orgânica a partir da fixação de carbono inorgânico. Estes seres podem realizar quimiossíntese (são quimioautotróficos) ou fotossíntese (são fotoautotróficos).
- Heterotróficos, se, não conseguindo fixar carbono inorgânico, o extraem de compostos orgânicos. Consoante o modo como essa extracção ocorre, os heterotróficos são-no por ingestão com digestão extra ou intra-celular ou por absorção. Há, ainda, alguns destes seres que, para além destas fontes de energia orgânicas, a obtêm, sem fixação de C, a partir da luz (fotoheterotróficos) ou da oxidação de moléculas inorgânicas (quimioheterotróficos).
- Interacção trófica nos ecossistemas. Os seres autotróficos são produtores, enquanto os heterotróficos são consumidores de dois tipos: micro (heterotrofismo por absorção) ou macroconsumidores (heterotrofismo por ingestão).
- Composição bioquímica. Sendo um dos argumentos do evolucionismo, a análise bioquímica permite estabelecer relações entre os seres e, por isso, classificá-los.
13.1. Classificação taxonómica actual
Desde um sistema de classificação com dois grupos de seres vivos até um sistema com cinco, a taxonomia evoluiu significativamente.

Actualmente, utiliza-se o sistema de classificação de Whittaker modificado que agrupa os seres vivos em cinco largos grupos – os reinos Monera, Protista, Plantae, Fungi e Animalia – que se distinguem em função do nível de organização celular, tipo de nutrição e interacções nos ecossistemas dos organismos que neles figuram, como pode observar-se no seguinte quadro (retirado daqui).

O reino é o taxon (= categoria taxonómica de plural taxa) mais abrangente da classificação taxonómica hierárquica que usamos, estando subdividido em filos, que por sua vez contêm várias classes. Cada classe engloba várias ordens, que incluem várias famílias, nas quais se inserem vários géneros e é dentro destes grupos que estão as espécies. Destas sete taxa principais (reino, filo, classe, ordem, família, género e espécie)viii, todas foram criadas pelo Homem, com excepção da espécie que, definindo-se como um conjunto de indivíduos que partilha o mesmo fundo genético e pode, por isso, cruzar-se entre si, originando descendência fértil, é um grupo natural (= delimitado pela própria natureza).
De acordo com este mesmo sistema, todas as categorias taxonómicas são palavras latinas ou latinizadas, surgindo, por isso, em itálico ou sublinhadas. Em relação às famílias, deve notar-se que, geralmente, dentro do reino Animalia, todas terminam em –idae, e, nas plantas, em –aceae.
A nomenclatura das espécies destaca-se devido ao facto de ser, ao contrário das outras 6 taxa principais, que se resumem a uma só palavra, binominal: o nome da espécie inclui o nome do género, com inicial maiúscula, e o restritivo ou epíteto específico, totalmente em letras minúsculas (por exemplo, para o lobo, Canis é o género e lupus é o epíteto específico, sendo o nome da espécie Canis lupus). Similarmente, as subespécies têm uma nomenclatura trinominal que obedece à fórmula género + restritivo específico + restritivo subespecífico (por exemplo, Homo sapiens sapiens). Nomes de espécies e subespécies são, geralmente, acompanhado por uma abreviatura do nome do taxonomista que catalogou em primeiro lugar os indivíduos desse tipo e, após uma vírgula, a data de publicação da descoberta.
13.3. Classificação em domínios
Em 1970, através de estudos baseados em critérios moleculares e, mais especificamente, nas sequências nucleotídicas de RNA ribossómico de vários tipos de procariontes, Carl Woese concluiu que existiam dois grupos distintos destes – arqueobactérias e eubactérias, sendo que as primeiras divergiram em primeiro lugar do ancestral comum que deu origem a ambos os grupos – o que o levou a propor uma classificação baseada em três domínios: Archaea (a que corresponde o reino Archeobacteria) Bacteria (a que corresponde o reino Eubacteria) e Eukarya (a que correspondem os reinos Protista, Plantae, Fungi e Animalia).

 i Do sangue que chega ao nível dos tecidos, cerca de 99% do volume retorna ao sistema venoso. Os restantes 1% vão formar a linfa intersticial, que é um líquido composto por plasma e leucócitos que actua como intermediário das trocas entre o sangue e os tecidos. Este fluido é, depois, recolhido por vasos capilares linfáticos, passando a designar-se linfa circulante, que se reúnem em canais colectores até atingirem o canal torácico (que conduz a linfa de volta à corrente sanguínea através da veia subclávia esquerda) ou o canal linfático direito (que conduz a linfa de volta à corrente sanguínea através da veia subclávia esquerda). A circulação da linfa deve-se à à pressão osmótica do sangue e à pressão pela contracção dos músculos. Ao longo dos vasos linfáticos surgem os gânglios linfáticos, onde há grandes quantidades de linfócitos (um tipo de leucócitos). Resumindo, o sistema linfático (conjunto gânglios + vasos linfáticos) mantém o equilíbrio dos iões no corpo, transporta certos ácidos gordos das vilosidades intestinais para o sangue, contribui para a imunidade do organismo e assegura a entrada da linfa no sistema circulatório. ↑
i Do sangue que chega ao nível dos tecidos, cerca de 99% do volume retorna ao sistema venoso. Os restantes 1% vão formar a linfa intersticial, que é um líquido composto por plasma e leucócitos que actua como intermediário das trocas entre o sangue e os tecidos. Este fluido é, depois, recolhido por vasos capilares linfáticos, passando a designar-se linfa circulante, que se reúnem em canais colectores até atingirem o canal torácico (que conduz a linfa de volta à corrente sanguínea através da veia subclávia esquerda) ou o canal linfático direito (que conduz a linfa de volta à corrente sanguínea através da veia subclávia esquerda). A circulação da linfa deve-se à à pressão osmótica do sangue e à pressão pela contracção dos músculos. Ao longo dos vasos linfáticos surgem os gânglios linfáticos, onde há grandes quantidades de linfócitos (um tipo de leucócitos). Resumindo, o sistema linfático (conjunto gânglios + vasos linfáticos) mantém o equilíbrio dos iões no corpo, transporta certos ácidos gordos das vilosidades intestinais para o sangue, contribui para a imunidade do organismo e assegura a entrada da linfa no sistema circulatório. ↑ii A bainha de mielina, isolante, impede a alteração do potencial da membrana que lhe subjaz, obrigando o impulso eléctrico a “saltar” de uma zona não mielinizada para a seguinte (ou seja, entre os nódulos de Ranvier) e, deste modo, acelerando, como previamente explicado, a sua velocidade de propagação.) ↑
iii A intensidade do estímulo tem que ultrapassar o limiar de excitabilidade do neurónio, mas, desde que o exceda, desencadeará um mesmo potencial de acção, independentemente do seu valor. ↑
iv Existem limites para a osmoconformidade, visto que condições extremas de salinidade (concentrações extremamente altas ou extremamente baixas de sais) podem levar à desnaturação das proteínas essenciais ao funcionamento do ser ou à plasmólise ou turgescência das suas células e, como consequência, à sua morte. Pode, por isso, concluir-se que a salinidade é um factor limitante. ↑
v Mesmo que os gâmetas se formem por mitoses, são divisões de células que têm a sua originem noutras formadas por meiose pós-zigótica ou pré-espórica, como se analisará no âmbito dos ciclos de vida, pelo que conservam as características destas. ↑
vi A conjugação é um processo semelhante à fecundação, mas no qual os gâmetas intervenientes são indiferenciados. Por exemplo, no caso da espirogira, não existindo gâmetas femininos e masculinos, admite-se, por convenção, que o conteúdo celular que se movimenta é o gâmeta dador e o que permanece imóvel é o gâmeta receptor. ↑

viii As categorias intermédias são sinalizadas com prefixos como super, infra e sub. ↑

4 comentários:
Esse blog é maravilhoso. Muito organizado e completo. Obrigada por existir, salvaram meus estudos!
TRABALHO MARAVILHOSO-. PARABÉNS
Excelente Blog!
Melhor forma de organização e disponibilização do conteúdo que encontrei.
Acredito que houve algum engano na seção dos sistemas circulatórios, onde é descrito que o sangue venoso que chega ao átrio (aurícula) direito passa ao ventrículo direito através da válvula BICÚSPIDE (mitral). Não seria a válvula TRICÚSPIDE?
Excelente acervo!
(link: http://webpages.fc.ul.pt/~rfcruz/relats/imagens/heart.jpg )
Enviar um comentário